

Parvalbumin-Positive Inhibitory Interneurons Oppose Propagation But Favor Generation of Focal Epileptiform Activity
- PMID: 26134638
- PMCID: PMC6605139
- DOI: 10.1523/JNEUROSCI.5117-14.2015
Parvalbumin-Positive Inhibitory Interneurons Oppose Propagation But Favor Generation of Focal Epileptiform Activity
Abstract
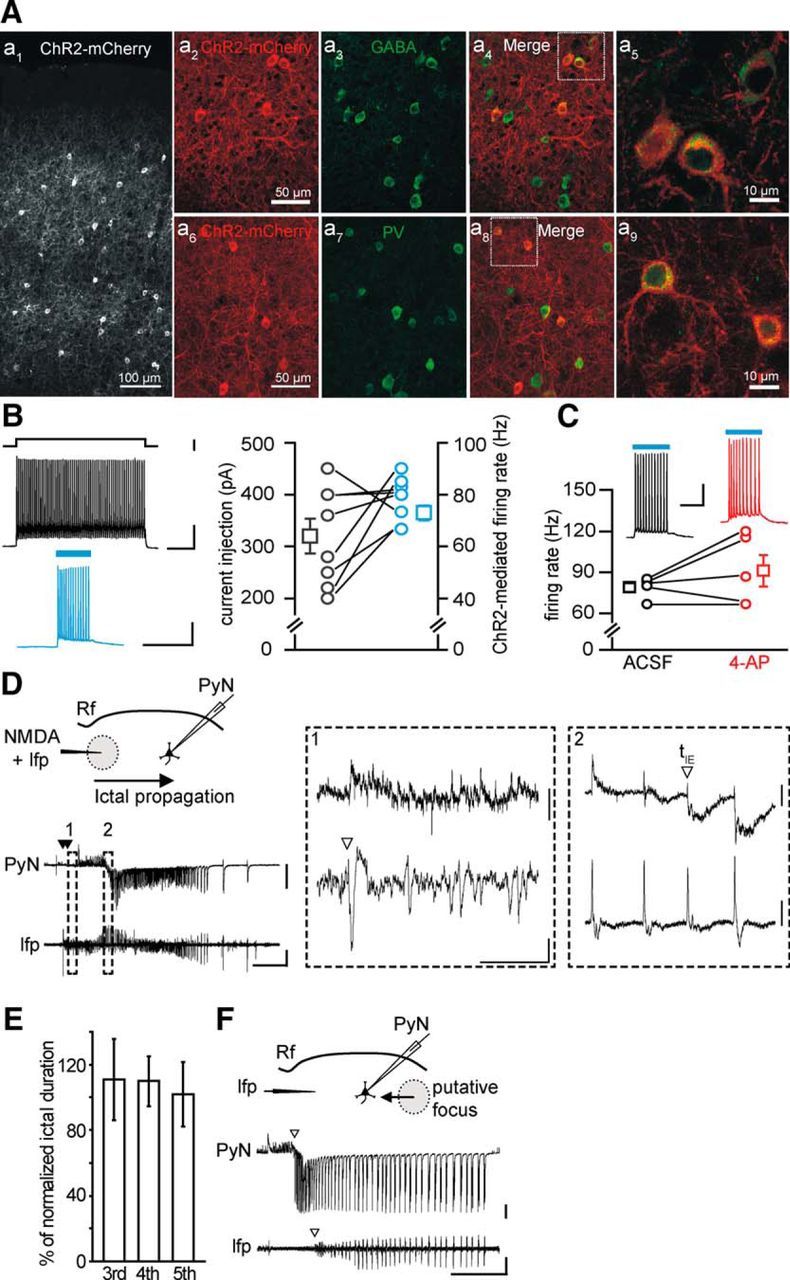
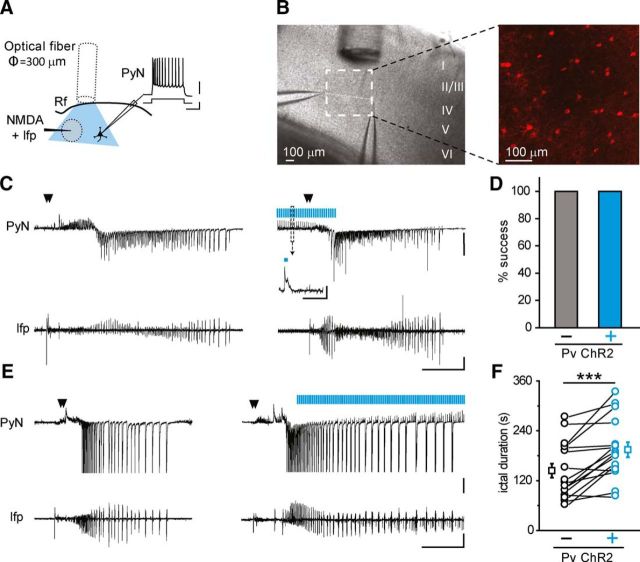
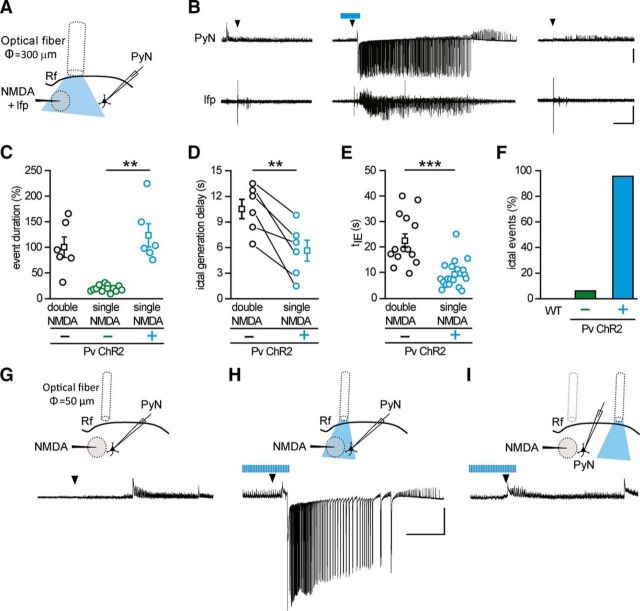
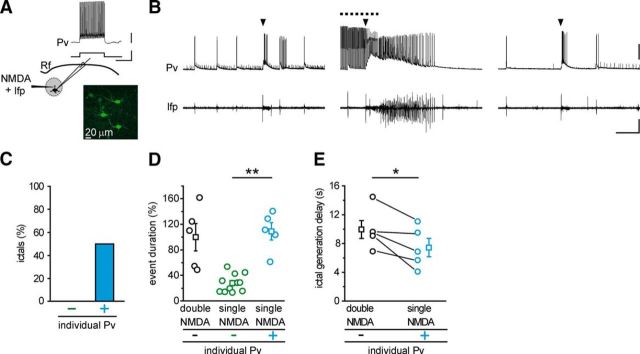
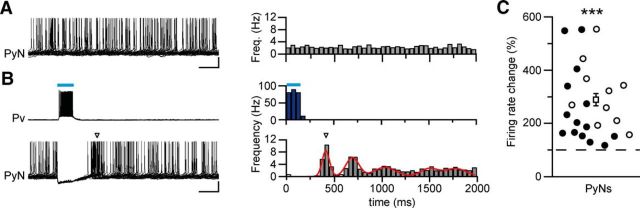
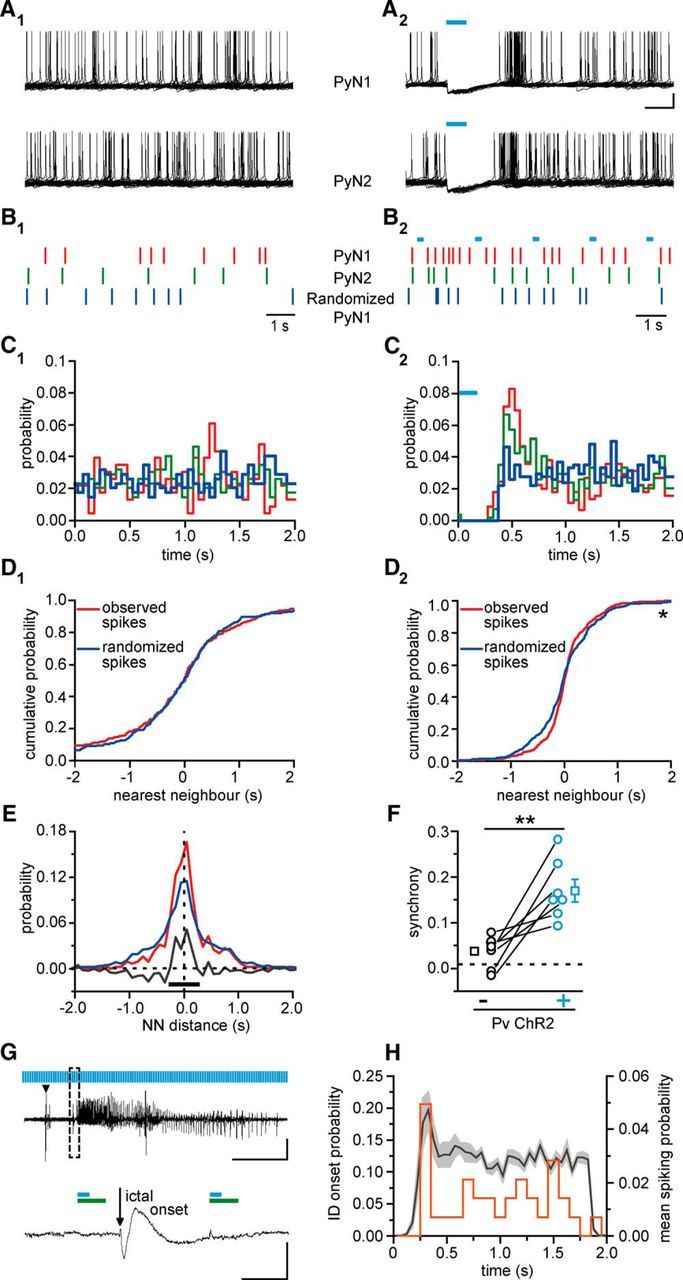
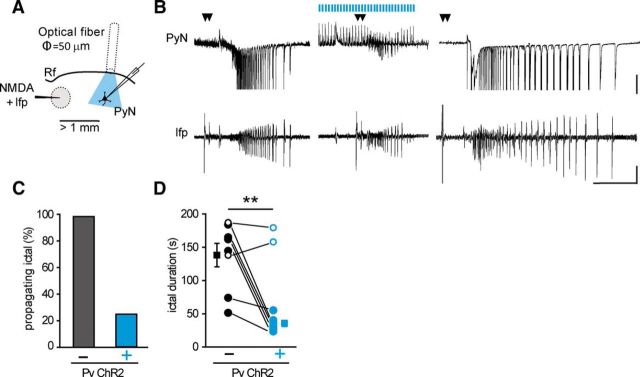
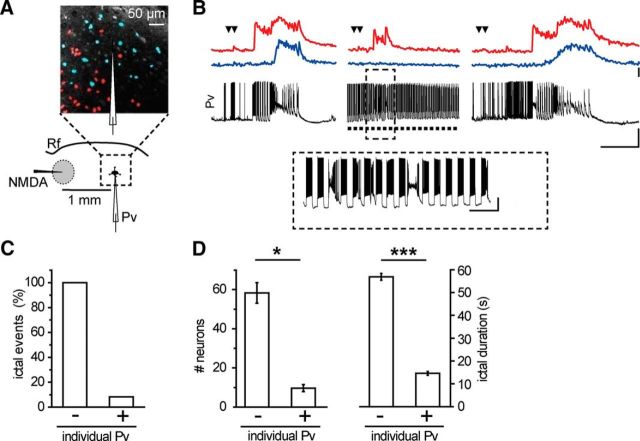
Parvalbumin (Pv)-positive inhibitory interneurons effectively control network excitability, and their optogenetic activation has been reported to block epileptic seizures. An intense activity in GABAergic interneurons, including Pv interneurons, before seizures has been described in different experimental models of epilepsy, raising the hypothesis that an increased GABAergic inhibitory signal may, under certain conditions, initiate seizures. It is therefore unclear whether the activity of Pv interneurons enhances or opposes epileptiform activities. Here we use a mouse cortical slice model of focal epilepsy in which the epileptogenic focus can be identified and the role of Pv interneurons in the generation and propagation of seizure-like ictal events is accurately analyzed by a combination of optogenetic, electrophysiological, and imaging techniques. We found that a selective activation of Pv interneurons at the focus failed to block ictal generation and induced postinhibitory rebound spiking in pyramidal neurons, enhancing neuronal synchrony and promoting ictal generation. In contrast, a selective activation of Pv interneurons distant from the focus blocked ictal propagation and shortened ictal duration at the focus. We revealed that the reduced ictal duration was a direct consequence of the ictal propagation block, probably by preventing newly generated afterdischarges to travel backwards to the original focus of ictal initiation. Similar results were obtained upon individual Pv interneuron activation by intracellular depolarizing current pulses. The functional dichotomy of Pv interneurons here described opens new perspectives to our understanding of how local inhibitory circuits govern generation and spread of focal epileptiform activities.
Keywords: channelrhodopsin-2; cortex; epileptiform activity; optogenetics; parvalbumin; rebound spiking.
Copyright © 2015 the authors 0270-6474/15/359544-14$15.00/0.
Figures
Similar articles
-
Disrupting Epileptiform Activity by Preventing Parvalbumin Interneuron Depolarization Block.J Neurosci. 2021 Nov 10;41(45):9452-9465. doi: 10.1523/JNEUROSCI.1002-20.2021. Epub 2021 Oct 5. J Neurosci. 2021. PMID: 34611025 Free PMC article.
-
Parvalbumin-expressing inhibitory interneurons in auditory cortex are well-tuned for frequency.J Neurosci. 2013 Aug 21;33(34):13713-23. doi: 10.1523/JNEUROSCI.0663-13.2013. J Neurosci. 2013. PMID: 23966693 Free PMC article.
-
Global optogenetic activation of inhibitory interneurons during epileptiform activity.J Neurosci. 2014 Feb 26;34(9):3364-77. doi: 10.1523/JNEUROSCI.2734-13.2014. J Neurosci. 2014. PMID: 24573293 Free PMC article.
-
GABAA-Receptor Signaling and Ionic Plasticity in the Generation and Spread of Seizures.In: Noebels JL, Avoli M, Rogawski MA, Vezzani A, Delgado-Escueta AV, editors. Jasper's Basic Mechanisms of the Epilepsies. 5th edition. New York: Oxford University Press; 2024. Chapter 6. In: Noebels JL, Avoli M, Rogawski MA, Vezzani A, Delgado-Escueta AV, editors. Jasper's Basic Mechanisms of the Epilepsies. 5th edition. New York: Oxford University Press; 2024. Chapter 6. PMID: 39637123 Free Books & Documents. Review.
-
Parvalbumin Interneuron Dysfunction in Neurological Disorders: Focus on Epilepsy and Alzheimer's Disease.Int J Mol Sci. 2024 May 19;25(10):5549. doi: 10.3390/ijms25105549. Int J Mol Sci. 2024. PMID: 38791587 Free PMC article. Review.
Cited by
-
Mesoscopic Mapping of Ictal Neurovascular Coupling in Awake Behaving Mice Using Optical Spectroscopy and Genetically Encoded Calcium Indicators.Front Neurosci. 2021 Jul 23;15:704834. doi: 10.3389/fnins.2021.704834. eCollection 2021. Front Neurosci. 2021. PMID: 34366781 Free PMC article.
-
Optogenetic activation of VGLUT2-expressing excitatory neurons blocks epileptic seizure-like activity in the mouse entorhinal cortex.Sci Rep. 2017 Feb 23;7:43230. doi: 10.1038/srep43230. Sci Rep. 2017. PMID: 28230208 Free PMC article.
-
Parvalbumin Role in Epilepsy and Psychiatric Comorbidities: From Mechanism to Intervention.Front Integr Neurosci. 2022 Feb 17;16:765324. doi: 10.3389/fnint.2022.765324. eCollection 2022. Front Integr Neurosci. 2022. PMID: 35250498 Free PMC article. Review.
-
Reduced Cholecystokinin-Expressing Interneuron Input Contributes to Disinhibition of the Hippocampal CA2 Region in a Mouse Model of Temporal Lobe Epilepsy.J Neurosci. 2023 Oct 11;43(41):6930-6949. doi: 10.1523/JNEUROSCI.2091-22.2023. Epub 2023 Aug 29. J Neurosci. 2023. PMID: 37643861 Free PMC article.
-
Seizing Control: From Current Treatments to Optogenetic Interventions in Epilepsy.Neuroscientist. 2017 Feb;23(1):68-81. doi: 10.1177/1073858415619600. Epub 2016 Jul 9. Neuroscientist. 2017. PMID: 26700888 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials