

Mitogen-activated protein kinase kinase 5 (MKK5)-mediated signalling cascade regulates expression of iron superoxide dismutase gene in Arabidopsis under salinity stress
- PMID: 26136265
- PMCID: PMC4566985
- DOI: 10.1093/jxb/erv305
Mitogen-activated protein kinase kinase 5 (MKK5)-mediated signalling cascade regulates expression of iron superoxide dismutase gene in Arabidopsis under salinity stress
Erratum in
-
Corrigendum to: Mitogen-activated protein kinase kinase 5 (MKK5)-mediated signaling cascade regulates expression of iron superoxide dismutase gene in Arabidopsis under salinity stress.J Exp Bot. 2021 Jun 22;72(13):5094. doi: 10.1093/jxb/erab133. J Exp Bot. 2021. PMID: 33956081 Free PMC article. No abstract available.
-
Correction.J Exp Bot. 2022 Nov 2;73(19):6985. doi: 10.1093/jxb/erac332. J Exp Bot. 2022. PMID: 36102817 Free PMC article. No abstract available.
Abstract
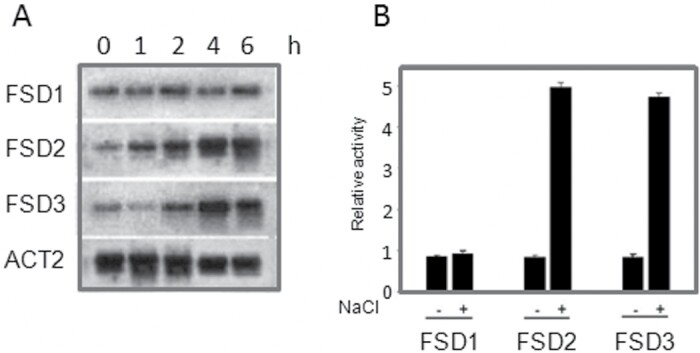
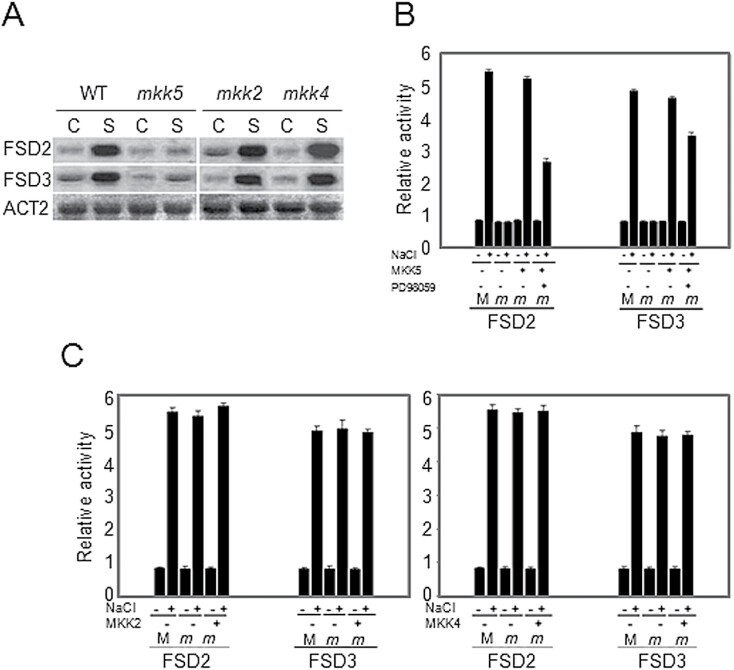
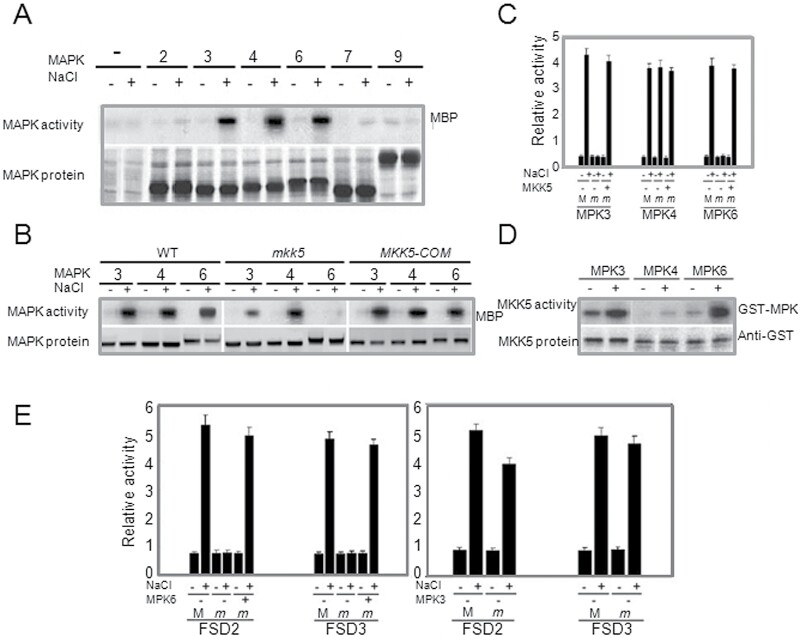
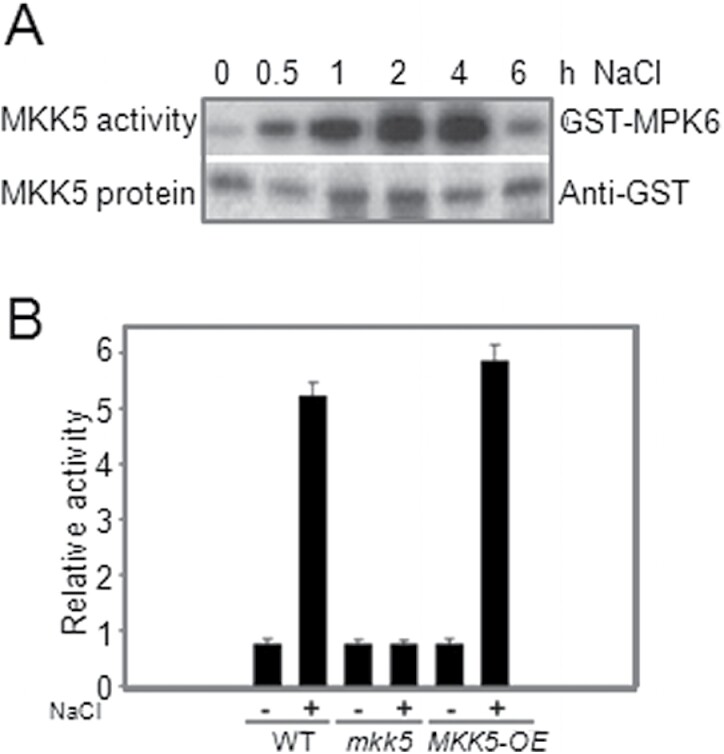
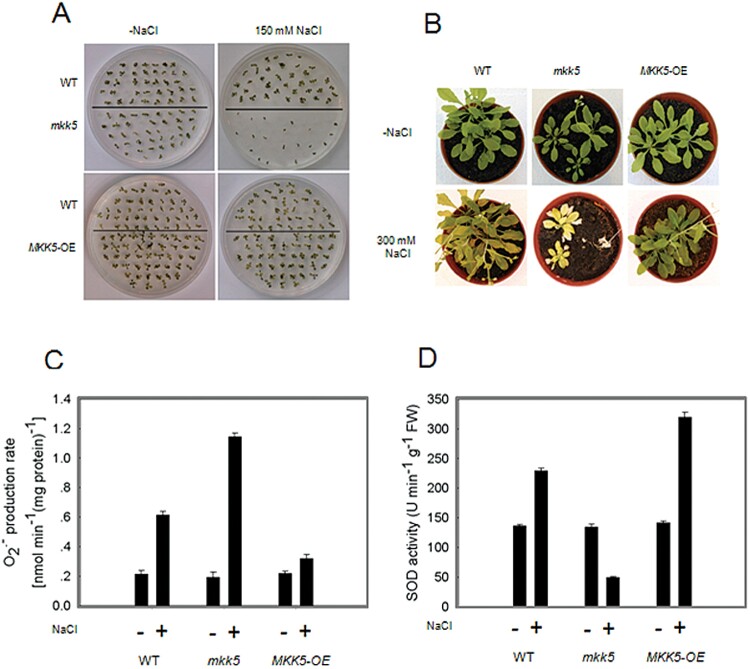
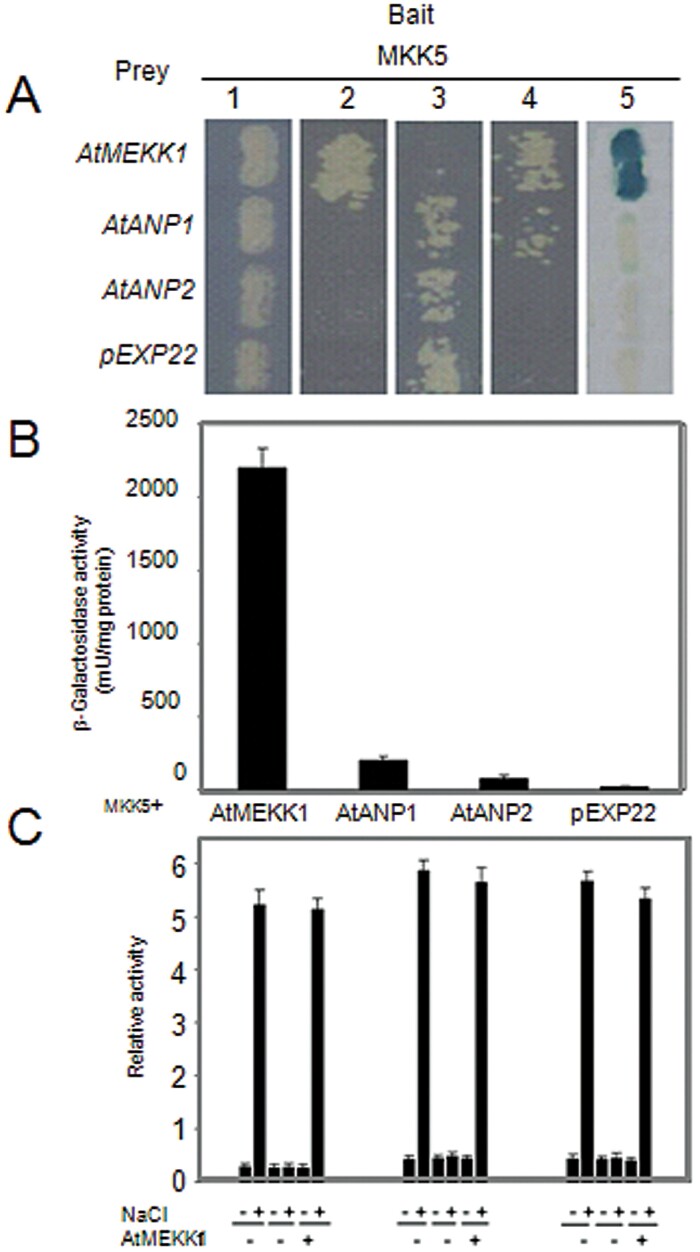
Superoxide dismutases (SODs) are involved in plant adaptive responses to biotic and abiotic stresses but the upstream signalling process that modulates their expression is not clear. Expression of two iron SODs, FSD2 and FSD3, was significantly increased in Arabidopsis in response to NaCl treatment but blocked in transgenic MKK5-RNAi plant, mkk5. Using an assay system for transient expression in protoplasts, it was found that mitogen-activated protein kinase kinase 5 (MKK5) was also activated in response to salt stress. Overexpression of MKK5 in wild-type plants enhanced their tolerance to salt treatments, while mkk5 mutant exhibited hypersensitivity to salt stress in germination on salt-containing media. Moreover, another kinase, MPK6, was also involved in the MKK5-mediated iron superoxide dismutase (FSD) signalling pathway in salt stress. The kinase activity of MPK6 was totally turned off in mkk5, whereas the activity of MPK3 was only partially blocked. MKK5 interacted with the MEKK1 protein that was also involved in the salt-induced FSD signalling pathway. These data suggest that salt-induced FSD2 and FSD3 expressions are influenced by MEKK1 via MKK5-MPK6-coupled signalling. This MAP kinase cascade (MEKK1, MKK5, and MPK6) mediates the salt-induced expression of iron superoxide dismutases.
Keywords: Arabidopsis; FSD; iron superoxide dismutase; mitogen-activated protein kinase 6; mitogen-activated protein kinase kinase 5; salt stress..
© The Author 2015. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
Similar articles
-
MKK5 regulates high light-induced gene expression of Cu/Zn superoxide dismutase 1 and 2 in Arabidopsis.Plant Cell Physiol. 2013 Jul;54(7):1217-27. doi: 10.1093/pcp/pct072. Epub 2013 May 14. Plant Cell Physiol. 2013. PMID: 23677921
-
AIK1, A Mitogen-Activated Protein Kinase, Modulates Abscisic Acid Responses through the MKK5-MPK6 Kinase Cascade.Plant Physiol. 2017 Feb;173(2):1391-1408. doi: 10.1104/pp.16.01386. Epub 2016 Dec 2. Plant Physiol. 2017. PMID: 27913741 Free PMC article.
-
The heat shock factor A4A confers salt tolerance and is regulated by oxidative stress and the mitogen-activated protein kinases MPK3 and MPK6.Plant Physiol. 2014 May;165(1):319-34. doi: 10.1104/pp.114.237891. Epub 2014 Mar 27. Plant Physiol. 2014. PMID: 24676858 Free PMC article.
-
Suppression of MKK5 reduces ozone-induced signal transmission to both MPK3 and MPK6 and confers increased ozone sensitivity in Arabidopsis thaliana.Plant Signal Behav. 2009 Aug;4(8):687-92. doi: 10.4161/psb.4.8.9298. Epub 2009 Aug 3. Plant Signal Behav. 2009. PMID: 19820329 Free PMC article.
-
A close-up of regulatory networks and signaling pathways of MKK5 in biotic and abiotic stresses.Crit Rev Biotechnol. 2025 Mar;45(2):473-490. doi: 10.1080/07388551.2024.2344584. Epub 2024 May 26. Crit Rev Biotechnol. 2025. PMID: 38797669 Review.
Cited by
-
Molecular Regulation of Host Defense Responses Mediated by Biological Anti-TMV Agent Ningnanmycin.Viruses. 2019 Sep 3;11(9):815. doi: 10.3390/v11090815. Viruses. 2019. PMID: 31484426 Free PMC article.
-
CRK41 Modulates Microtubule Depolymerization in Response to Salt Stress in Arabidopsis.Plants (Basel). 2023 Mar 12;12(6):1285. doi: 10.3390/plants12061285. Plants (Basel). 2023. PMID: 36986973 Free PMC article.
-
The ABCISIC ACID INSENSITIVE (ABI) 4 Transcription Factor Is Stabilized by Stress, ABA and Phosphorylation.Plants (Basel). 2022 Aug 22;11(16):2179. doi: 10.3390/plants11162179. Plants (Basel). 2022. PMID: 36015481 Free PMC article.
-
Comparative phenotypic, physiological, and transcriptomic responses to drought and recovery in two Fraxinus species.BMC Plant Biol. 2025 Mar 18;25(1):348. doi: 10.1186/s12870-025-06372-6. BMC Plant Biol. 2025. PMID: 40098103 Free PMC article.
-
Traversing the Links between Heavy Metal Stress and Plant Signaling.Front Plant Sci. 2018 Feb 5;9:12. doi: 10.3389/fpls.2018.00012. eCollection 2018. Front Plant Sci. 2018. PMID: 29459874 Free PMC article. Review.
References
-
- Abel S, Theologis A. 1994. Transient transformation of Arabidopsis leaf protoplasts: a versatile experimental system to study gene expression. The Plant Journal 5, 421–428.. - PubMed
-
- Acharya BR, Jeon BW, Zhang W, Assmann SM. 2013. Open stomata (OST1) is limiting in abscisic acid responses of Arabidopsis guard cells. New Phytologist 200, 1049–1063.. - PubMed
-
- Andreasson E, Ellis B. 2010. Convergence and specificity in the Arabidopsis MAPK nexus. Trends in Plant Science 15, 106–113.. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
