

Review
doi: 10.1152/physiol.00007.2015.
Humans In Hypoxia: A Conspiracy Of Maladaptation?!
Affiliations
- PMID: 26136544
- PMCID: PMC4491526
- DOI: 10.1152/physiol.00007.2015
Item in Clipboard
Review
Humans In Hypoxia: A Conspiracy Of Maladaptation?!
Physiology (Bethesda).
2015 Jul.
Abstract
We address adaptive vs. maladaptive responses to hypoxemia in healthy humans and hypoxic-tolerant species during wakefulness, sleep, and exercise. Types of hypoxemia discussed include short-term and life-long residence at high altitudes, the intermittent hypoxemia attending sleep apnea, or training regimens prescribed for endurance athletes. We propose that hypoxia presents an insult to O2 transport, which is poorly tolerated in most humans because of the physiological cost.
©2015 Int. Union Physiol. Sci./Am. Physiol. Soc.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the author(s).
Figures
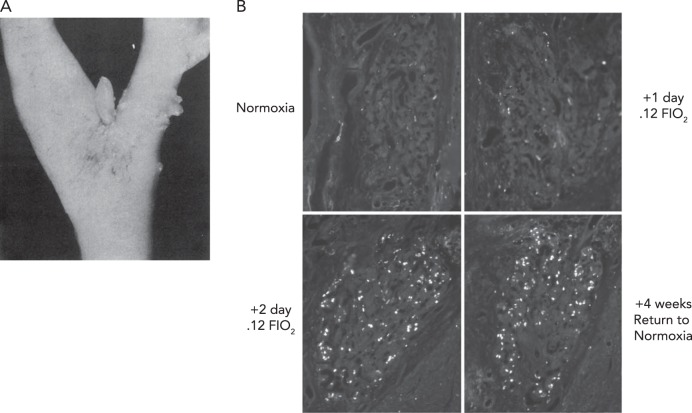
Human carotid chemoreceptor and immunofluorescence staining of bromodeoxyuridine A: human carotid chemoreceptor, located at the bifurcation of the common carotid artery. In health and in normoxia, the carotid body is usually 1–7 mm in length and 1–2 mm in diameter. The carotid chemoreceptor is located ∼10- to 15-s circulation time from the lung (under resting conditions), has sensory output via the carotid sinus nerve to the petrosal ganglion, with central projections via the nucleus of the solitary tract to the medullary pattern generation neurons, central chemoreceptor cells of the retrotrapezoid nucleus, and the paraventricular nucleus. Figure reproduced from Ref. with permission from BMJ Publishing Group, Ltd. B: immunofluorescence staining of bromodeoxyuridine (BrdU), a uridine analog that is stably incorporated into cells undergoing DNA synthesis, in carotid body type I cells from rats maintained in normoxia, exposed to sustained normobaric hypoxia (Fi O2, 0.12) for 1 (A) and 2 days, and rats exposed to hypoxia for 7 days, then returned to normoxia for another 30 days. Sustained hypoxia caused glomus cell proliferation that occurred mainly in the first 3 days of exposure. The new glomus cells survived for at least 30 days after return to normoxia. Scale bar, 50 μm. Figure reproduced from Ref. with permission.
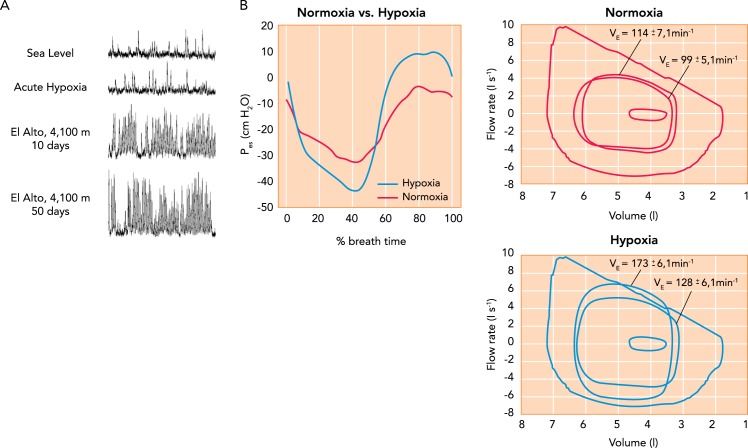
Hypoxic effects on muscle sympathetic nerve activity and on exercise ventilation and work of breathing A: time-dependent increase in muscle sympathetic nerve activity (MSNA) in a healthy human during sojourn at 4,100-m altitude. MSNA frequency remained elevated for up to 5 days following return to sea level. Figure reproduced with permission from M. Sander [also see Hansen et al. (45)]. B: effects on ventilation, expiratory flow limitation, and inspiratory and expiratory esophageal pressures (Pes) during constant load, heavy-intensity cycle exercise at 273 W in normoxia (95% SaO2, 69% of peak work rate) vs. acute hypoxia (Fi O2, 15; SaO2, ∼81%; 99% of peak work rate) in trained cyclists. At right, note the increased ventilation in hypoxia vs. normoxia at 3 min and at termination (9 min) of the same 273-W work rate. The maximum volitional flow-volume loop (obtained at rest) is the largest loop shown (inspiration down), and the three loops within this are average tidal breaths at rest and during exercise at 3 min and exercise termination. Note the occurrence of expiratory flow limitation in hypoxia near end-exercise. At left, negative Pes occurs during inspiration and positive Pes during expiration. These values for Pes were obtained at end-exercise in both normoxia and hypoxia. Ratings of perceived dyspnea on a 10-point scale at end exercise of equal durations averaged 4.4 ± 0.5 in normoxia and 8.7 ± 0.3 in acute hypoxia. Figure reproduced from Ref. with permission.
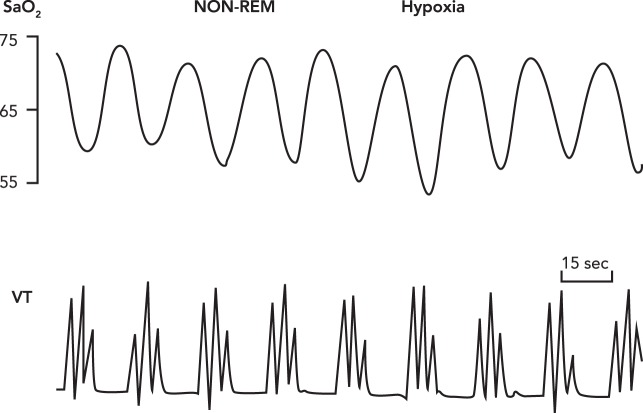
Periodic breathing with apneas Periodic breathing with apneas interspersed with three to four augmented tidal volumes occurring in 15- to 25-s cycles thought NREM sleep in a healthy sojourner to 4,100-m altitude. The “average” SaO2 is substantially reduced, and PaCO2 increased from wakefulness to NREM sleep in hypoxia (not shown), although little difference or even slight increases in the average SaO2 values between periodic and non-periodic patterns have been reported during sleep in hypoxia (14, 20, 43, 124). As explained in Adaptive IH, it is the oscillatory periods of HbO2 desaturation and resaturation that enhance the rate of free-radical production and their cardiovascular sequelae “. . . consistent with the notion that relevant enzymes are generating superoxide in response to the rate at which the electron receptor (O2) is introduced into the cell” (60). Reproduced from Ref. with permission from J Physiol.
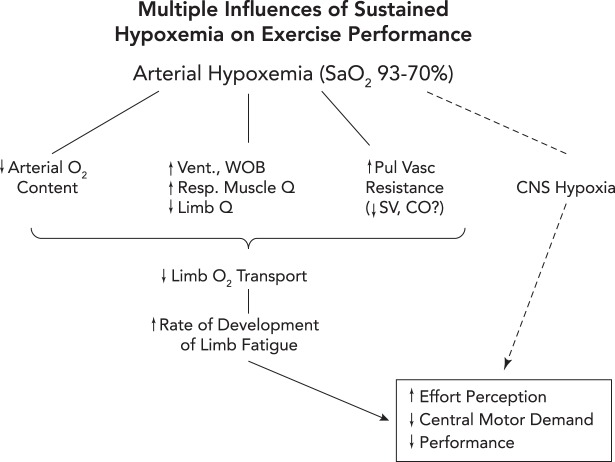
Three potential routes to exacerbation of exercise-induced central and peripheral (locomotor muscle) fatigue and reduction in exercise performance in hypoxia 1: reduced O2 transport to locomotor muscles via a) exercise-induced arterial hypoxemia (a); reductions in locomotor muscle blood flow secondary to an augmented sympathetic vasoconstriction of locomotor muscle vasculature elicited by high levels of respiratory muscle work and fatigue (b) (29); and/or stroke volume and cardiac output may be compromised via high left ventricular afterload induced by positive intrathoracic pressure on expiration in the face of airway expiratory flow limitation (70) (see also FIGURE 2A) or by increased pulmonary vascular resistance causing interventricular septal deviation toward the left ventricle, thereby altering left ventricular geometry and delaying filling (c) (56, 79). 2: CNS hypoxia, especially at extreme levels of arterial O2 desaturation (<75% SaO2), will inhibit central motor command (likely linked to reduced neurotransmitter turnover) and limit performance even in the face of limited amounts of peripheral muscle fatigue (6). 3: peripheral and central fatigue are causally linked via feedback inhibition of central motor output through group III–IV muscle afferents. When these afferents were partially blocked during a cycling time trial, the cyclist “chose” a much higher power output (vs. placebo control) in the initial half of the trial, leading to excessive muscle metabolite accumulation, reduced power output, and excessive locomotor muscle fatigue at end-exercise (5).
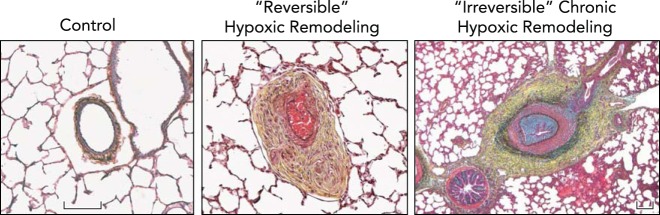
Chronic hypoxic exposure leads to severe pulmonary vascular remodeling, which is often reversible upon acute return to normoxia, but in extreme cases may be irreversible Distal pulmonary arteries of neonatal calves raised at ambient altitude (Control) are thin-walled and highly compliant, with minimal surrounding adventitia, whereas neonatal calves, exposed to 2 wk of hypobaric hypoxia, develop severe pulmonary hypertension, with mean PAP often exceeding 80 mmHg, and have pulmonary arteries that demonstrate marked fibro-proliferative changes in media and adventitia accompanied by formation of vasa vasorum (Reversible Hypoxic Remodeling). This profound remodeling, however, is reversible, and spontaneously gradually resolves when animals are returned to normobaric conditions. In older (5- to 9-mo-old) calves that were raised at moderately high altitudes (3,000-3,500 m), hypoxia-associated vascular remodeling and pulmonary hypertensive process can become severe and not regress in response to normoxia (Irreversible Chronic Hypoxic Remodeling). These changes are observed in animals exhibiting signs and symptoms of severe right heart failure. Remodeling of pulmonary arteries in these animals include severe neointimal, often occlusive, lesions as well as profound adventitial and medial thickening and fibrosis. Figure and legend were provided by Prof. Kurt Stenmark, University of Colorado, and used with his permission.
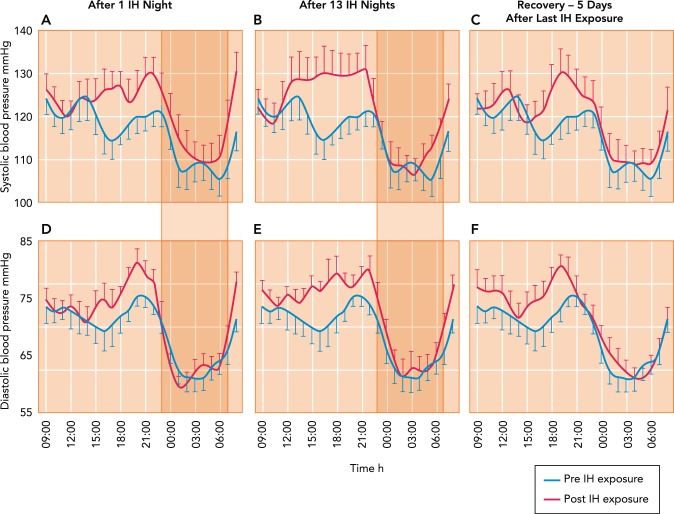
Effects on ambulatory systemic blood pressure of nocturnal intermittent, oscillating hypoxemia Hour-by-hour effects on ambulatory systemic blood pressure of nocturnal intermittent, oscillating hypoxemia consisting of 8 h of daily of oscillatory poikilocapnic hypoxia (Fi O2 of 0.13 every 105 s alternated with 15 s of Fi O2 of 1.0, producing periodic swings of Sp O2 of ∼88–98%) in healthy young adults. Note that increases in blood pressure resulting from IH occurred 1) just before waking (5–7 AM), then returned to normal until increasing again over the late afternoon and evening; 2) during the daytime hours after 1 night of IH, and increased further after 13 IH nights; and 3) returned to normal after 5 nights in normoxia. The IH effects on daytime BP were accompanied by significant increases in MSNA burst frequency and reductions in baroreceptor sensitivity. Figure was reproduced from Ref. with permission from the European Respiratory Society.
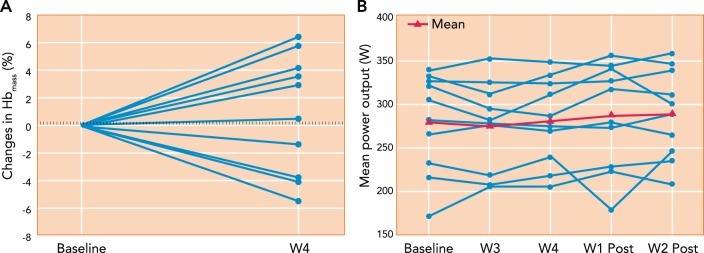
Effects of “live high:train low” training regimen in 10 endurance-trained cyclists on Hb mass and mean power output during a 26-km time trial The cyclists were exposed to daily normobaric hypoxia simulating 3,000-m altitude for 16 h/day for 28 nights and trained daily in normoxia. Note the high individual variability in the changes in Hb mass and mean power output during the time trials. The dashed lines represent the random error of the CO rebreathe test used for Hb mass. The group mean changes in Hb mass, time trial mean power output or performance, and V̇o 2 max (not shown) were not significantly different from those in a placebo group (n = 6) exposed to normoxia. Figure was reproduced from Ref. with permission.
References
-
- Ainslie PN, Lucas SJ, Burgess KR. Breathing and sleep at high altitude. Respir Physiol Neurobiol 188: 233–256, 2013. - PubMed
-
- Alexander JK, Grover RF. Mechanism of reduced cardiac stroke volume at high altitude. Clin Cardiol 6: 301–303, 1983. - PubMed
-
- Amann M, Pegelow DF, Jacques AJ, Dempsey JA. Inspiratory muscle work in acute hypoxia influences locomotor muscle fatigue and exercise performance of healthy humans. Am J Physiol Regul Integr Comp Physiol 293: R2036–R2045, 2007. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources