

The Influence of Gonadal Steroid Hormones on Immunoreactive Kisspeptin in the Preoptic Area and Arcuate Nucleus of Developing Agonadal Mice with a Genetic Disruption of Steroidogenic Factor 1
- PMID: 26138474
- PMCID: PMC4696913
- DOI: 10.1159/000437166
The Influence of Gonadal Steroid Hormones on Immunoreactive Kisspeptin in the Preoptic Area and Arcuate Nucleus of Developing Agonadal Mice with a Genetic Disruption of Steroidogenic Factor 1
Abstract
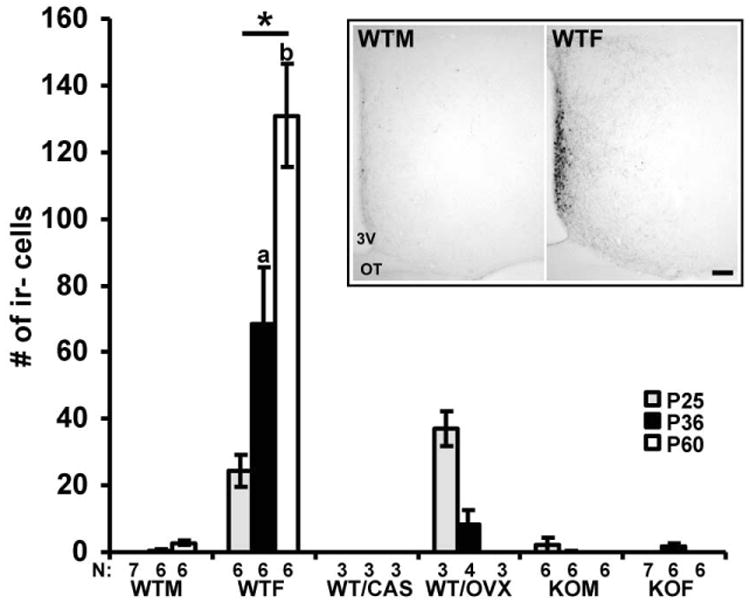
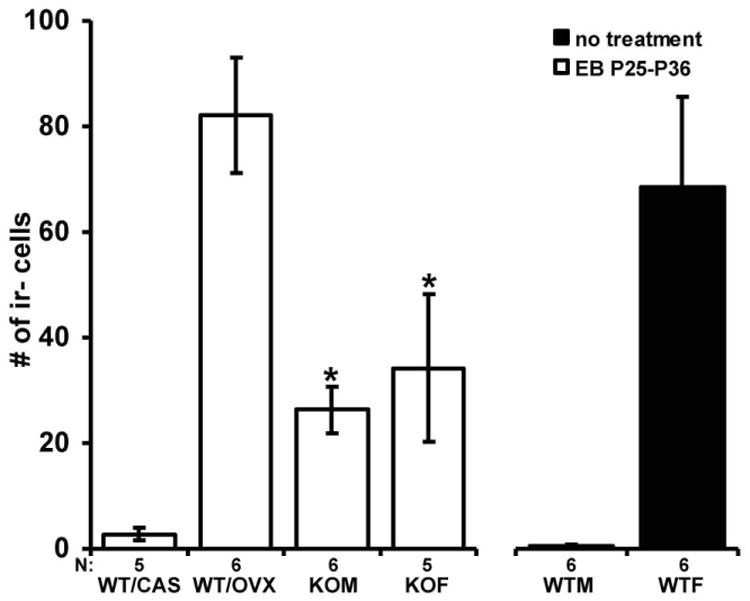
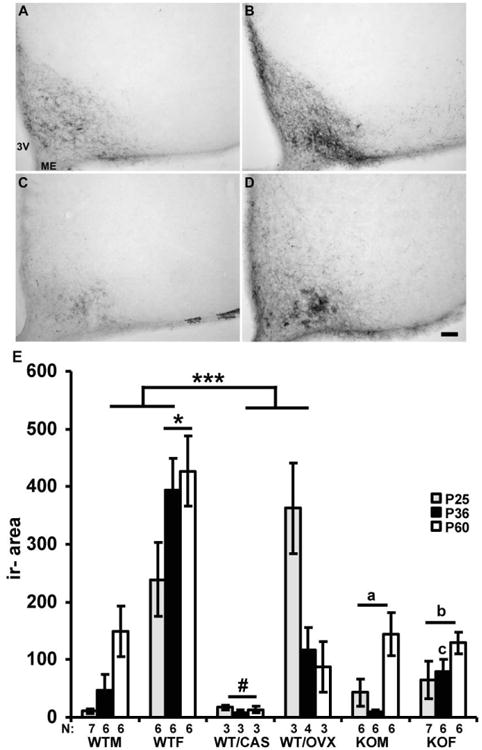
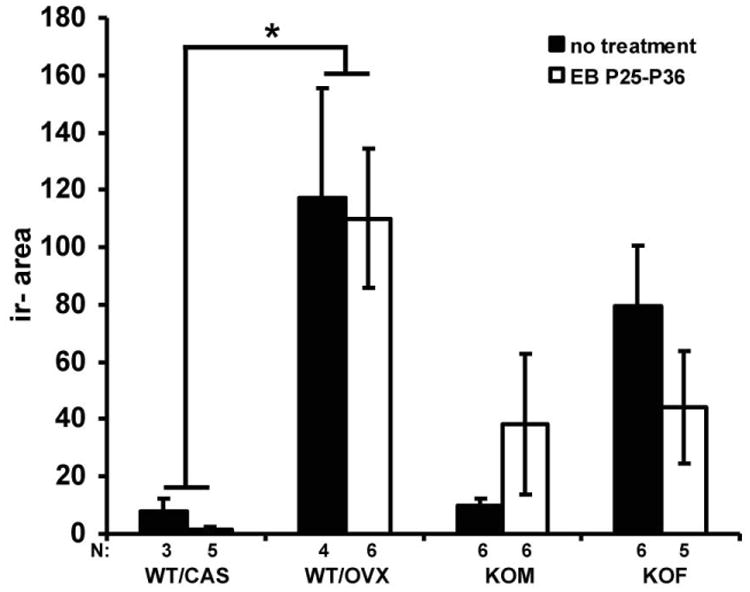
Kisspeptin, a regulator of reproductive function and puberty in mammals, is expressed in the rostral (anteroventral) periventricular nucleus (AVPV) and arcuate nucleus (Arc), and its expression is at least partially regulated by estradiol in rodents. The aim of the present study was to determine contributions of genetic factors and gonadal steroid hormones to the sexual differentiation of kisspeptin-immunoreactive (kisspeptin-ir) cell populations in the AVPV and Arc during postnatal development using agonadal steroidogenic factor 1 (SF-1) knockout (KO) mice. To examine the effects of gonadal hormones on pubertal development of kisspeptin neurons, SF-1 KO mice were treated with estradiol benzoate (EB) from postnatal day (P)25 to P36, and their brains were examined at P36. No sex differences were observed in the SF-1 KO mice during postnatal development and after treatment with EB - which failed to increase the number of kisspeptin-ir cells at P36 to the levels found in wild-type (WT) control females. This suggests that specific time periods of estradiol actions or other factors are needed for sexual differentiation of the pattern of immunoreactive kisspeptin in the AVPV. Kisspeptin immunoreactivity in the Arc was significantly higher in gonadally intact WT and SF-1 KO females than in male mice at P36 during puberty. Further, in WT and SF-1 KO females, but not in males, adult levels were reached at P36. This suggests that maturation of the kisspeptin system in the Arc differs between sexes and is regulated by gonad-independent mechanisms.
© 2015 S. Karger AG, Basel.
Figures
References
-
- Funes S, Hedrick JA, Vassileva G, Markowitz L, Abbondanzo S, Golovko A, Yang S, Monsma FJ, Gustafson EL. The KiSS-1 receptor GPR54 is essential for the development of the murine reproductive system. Biochem Biophys Res Commun. 2003;312:1357–1363. - PubMed
-
- Seminara SB, Messager S, Chatzidaki EE, Thresher RR, Acierno JS, Jr, Shagoury JK, Bo-Abbas Y, Kuohung W, Schwinof KM, Hendrick AG, Zahn D, Dixon J, Kaiser UB, Slaugenhaupt SA, Gusella JF, O'Rahilly S, Carlton MB, Crowley WF, Jr, Aparicio SA, Colledge WH. The GPR54 gene as a regulator of puberty. N Engl J Med. 2003;349:1614–1627. - PubMed
-
- Patterson M, Murphy KG, Thompson EL, Patel S, Ghatei MA, Bloom SR. Administration of kisspeptin-54 into discrete regions of the hypothalamus potently increases plasma luteinising hormone and testosterone in male adult rats. J Neuroendocrinol. 2006;18:349–354. - PubMed
-
- Plant TM, Ramaswamy S, Dipietro MJ. Repetitive activation of hypothalamic G protein-coupled receptor 54 with intravenous pulses of kisspeptin in the juvenile monkey (Macaca mulatta) elicits a sustained train of gonadotropin-releasing hormone discharges. Endocrinology. 2006;147:1007–1013. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
