

Bacterial microcompartments: widespread prokaryotic organelles for isolation and optimization of metabolic pathways
- PMID: 26148529
- PMCID: PMC4718714
- DOI: 10.1111/mmi.13117
Bacterial microcompartments: widespread prokaryotic organelles for isolation and optimization of metabolic pathways
Abstract
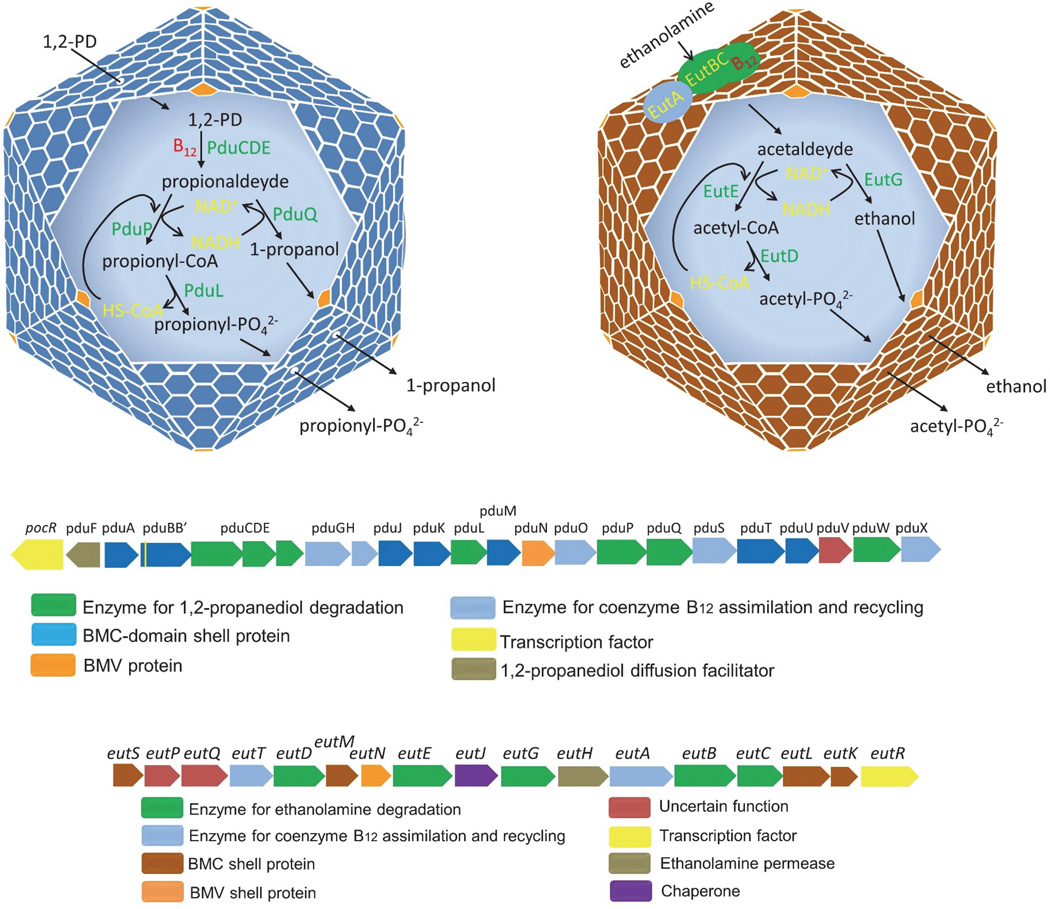
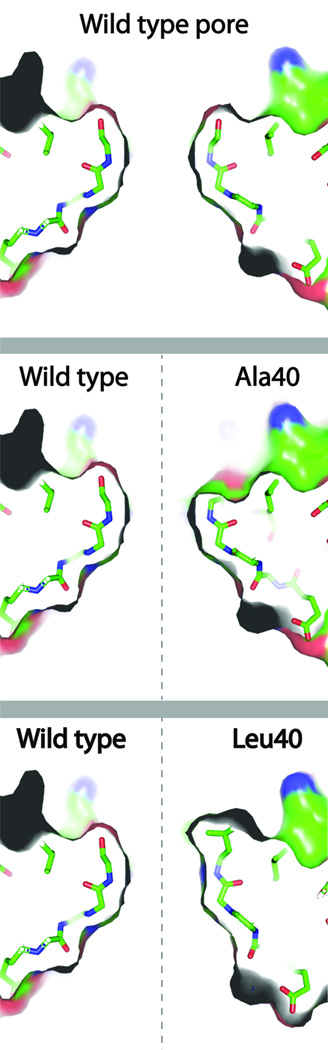
Prokaryotes use subcellular compartments for a variety of purposes. An intriguing example is a family of complex subcellular organelles known as bacterial microcompartments (MCPs). MCPs are widely distributed among bacteria and impact processes ranging from global carbon fixation to enteric pathogenesis. Overall, MCPs consist of metabolic enzymes encased within a protein shell, and their function is to optimize biochemical pathways by confining toxic or volatile metabolic intermediates. MCPs are fundamentally different from other organelles in having a complex protein shell rather than a lipid-based membrane as an outer barrier. This unusual feature raises basic questions about organelle assembly, protein targeting and metabolite transport. In this review, we discuss the three best-studied MCPs highlighting atomic-level models for shell assembly, targeting sequences that direct enzyme encapsulation, multivalent proteins that organize the lumen enzymes, the principles of metabolite movement across the shell, internal cofactor recycling, a potential system of allosteric regulation of metabolite transport and the mechanism and rationale behind the functional diversification of the proteins that form the shell. We also touch on some potential biotechnology applications of an unusual compartment designed by nature to optimize metabolic processes within a cellular context.
© 2015 John Wiley & Sons Ltd.
Figures



References
-
- Abdul-Rahman F, Petit E, Blanchard JL. The distribution of polyhedral bacterial microcompartments suggests frequent horizontal transfer and operon reassembly. J. Phylogen. Evolution. Biol. 2013;1
-
- Badger MR, Hanson D, Price GD. Evolution and diversity of CO2 concentrating mechanisms in cyanobacteria. Functional Plant Biology. 2002;29:161–173. - PubMed
-
- Baker SH, Williams DS, Aldrich HC, Gambrell AC, Shively JM. Identification and localization of the carboxysome peptide Csos3 and its corresponding gene in Thiobacillus neapolitanus. Arch Microbiol. 2000;173:278–283. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
