

Non-straight cell edges are important to invasion and engulfment as demonstrated by cell mechanics model
- PMID: 26148533
- PMCID: PMC4792343
- DOI: 10.1007/s10237-015-0697-6
Non-straight cell edges are important to invasion and engulfment as demonstrated by cell mechanics model
Abstract
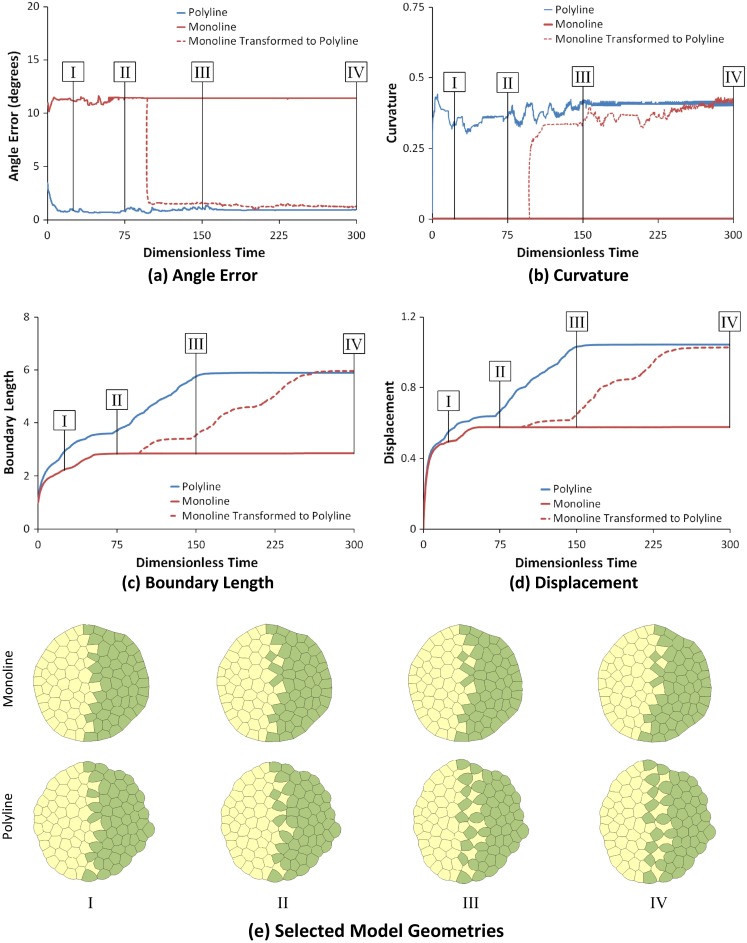
Computational models of cell-cell mechanical interactions typically simulate sorting and certain other motions well, but as demands on these models continue to grow, discrepancies between the cell shapes, contact angles and behaviours they predict and those that occur in real cells have come under increased scrutiny. To investigate whether these discrepancies are a direct result of the straight cell-cell edges generally assumed in these models, we developed a finite element model that approximates cell boundaries using polylines with an arbitrary number of segments. We then compared the predictions of otherwise identical polyline and monoline (straight-edge) models in a variety of scenarios, including annealing, single- and multi-cell engulfment, sorting, and two forms of mixing--invasion and checkerboard pattern formation. Keeping cell-cell edges straight influences cell motion, cell shape, contact angle, and boundary length, especially in cases where one cell type is pulled between or around cells of a different type, as in engulfment or invasion. These differences arise because monoline cells have restricted deformation modes. Polyline cells do not face these restrictions, and with as few as three segments per edge yielded realistic edge shapes and contact angle errors one-tenth of those produced by monoline models, making them considerably more suitable for situations where angles and shapes matter, such as validation of cellular force-inference techniques. The findings suggest that non-straight cell edges are important both in modelling and in nature.
Keywords: Cell engulfment; Cell mechanics; Cell mixing; Cell sorting; Cell–cell interactions; Checkerboard patterns; Computational modelling; Computer simulations; Finite element models; Invasion; Polyline models; Tissue engulfment.
Figures
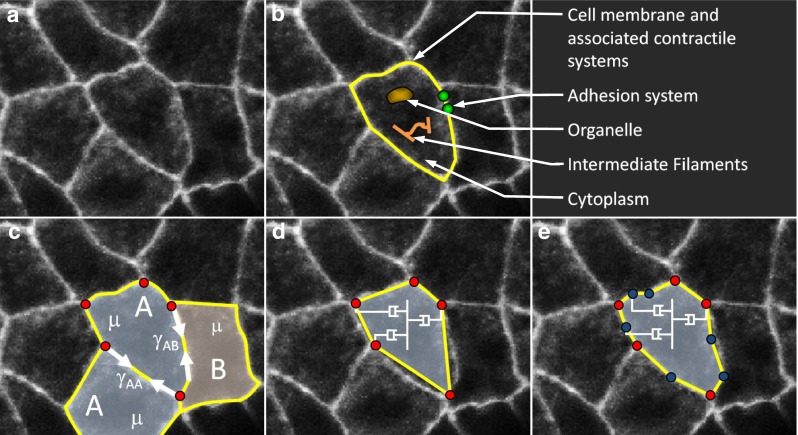
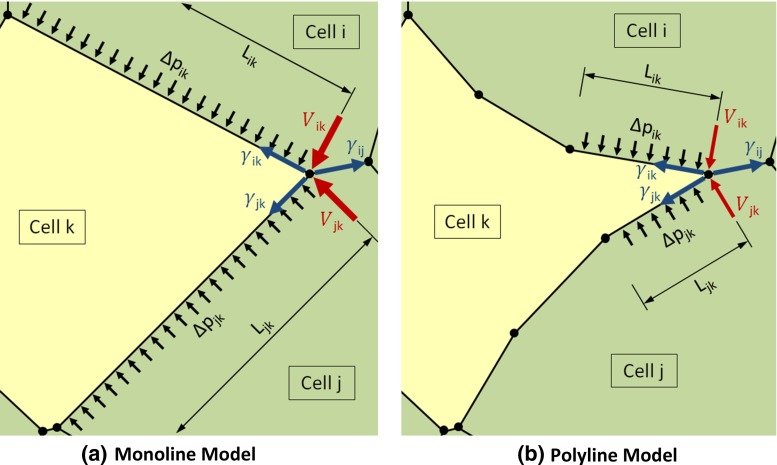
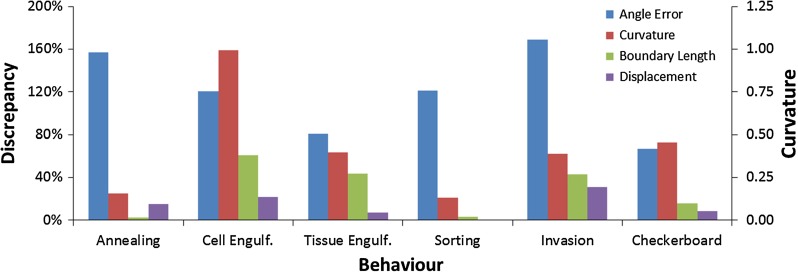
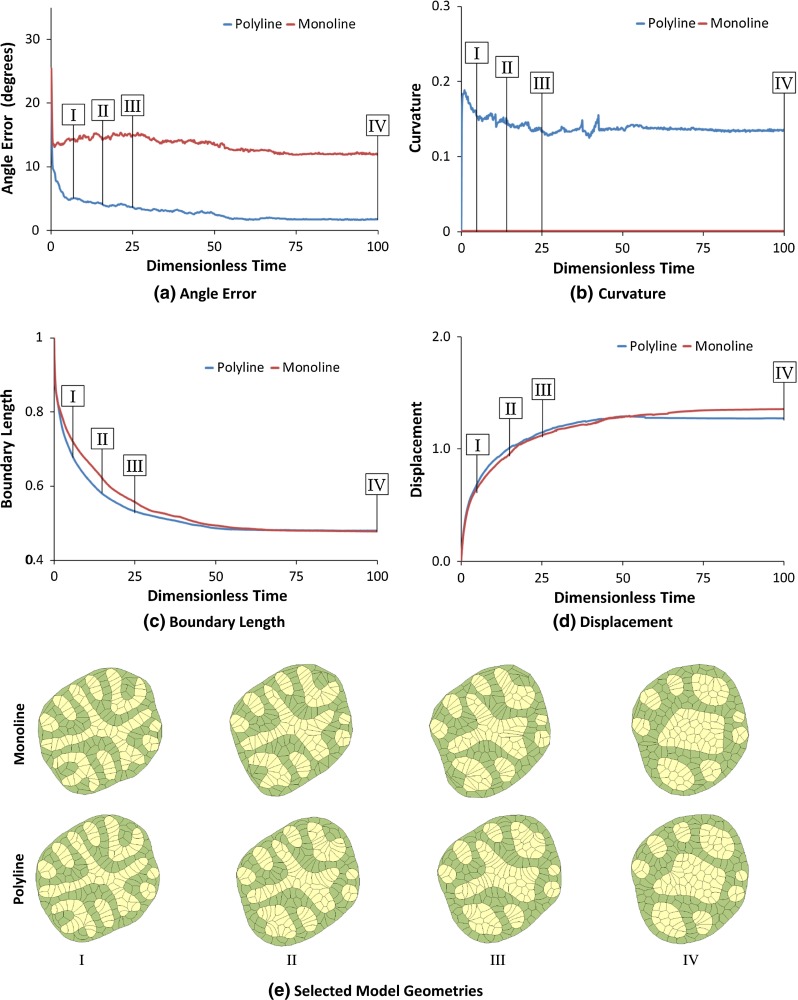
References
-
- Alber MS, Kiskowski M, Glazier JA, Jiang Y. On cellular automaton approaches to modeling biological cells. In: Rosenthal J, Gilliam D, editors. Mathematical systems theory in biology, communications, computation, and finance. New York: Springer; 2003. pp. 1–39.
-
- Armstrong PB, Armstrong MT. Intercellular invasion and the organizational stability of tissues: a role for fibronectin. Biochim Biophys Acta. 2000;1470:O9–20. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
