

Genome-Wide Screens in Saccharomyces cerevisiae Highlight a Role for Cardiolipin in Biogenesis of Mitochondrial Outer Membrane Multispan Proteins
- PMID: 26149385
- PMCID: PMC4539369
- DOI: 10.1128/MCB.00107-15
Genome-Wide Screens in Saccharomyces cerevisiae Highlight a Role for Cardiolipin in Biogenesis of Mitochondrial Outer Membrane Multispan Proteins
Abstract
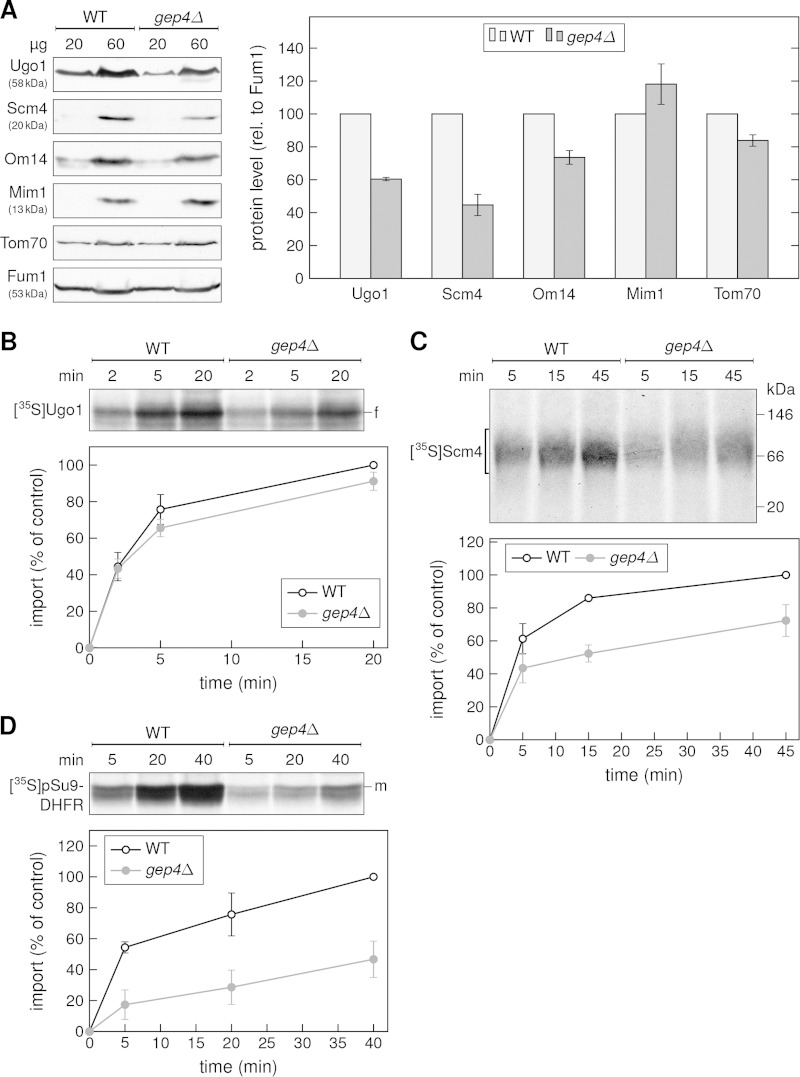
A special group of mitochondrial outer membrane (MOM) proteins spans the membrane several times via multiple helical segments. Such multispan proteins are synthesized on cytosolic ribosomes before their targeting to mitochondria and insertion into the MOM. Previous work recognized the import receptor Tom70 and the mitochondrial import (MIM) complex, both residents of the MOM, as required for optimal biogenesis of these proteins. However, their involvement is not sufficient to explain either the entire import pathway or its regulation. To identify additional factors that are involved in the biogenesis of MOM multispan proteins, we performed complementary high-throughput visual and growth screens in Saccharomyces cerevisiae. Cardiolipin (CL) synthase (Crd1) appeared as a candidate in both screens. Our results indeed demonstrate lower steady-state levels of the multispan proteins Ugo1, Scm4, and Om14 in mitochondria from crd1Δ cells. Importantly, MOM single-span proteins were not affected by this mutation. Furthermore, organelles lacking Crd1 had a lower in vitro capacity to import newly synthesized Ugo1 and Scm4 molecules. Crd1, which is located in the mitochondrial inner membrane, condenses phosphatidylglycerol together with CDP-diacylglycerol to obtain de novo synthesized CL molecules. Hence, our findings suggest that CL is an important component in the biogenesis of MOM multispan proteins.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures





References
-
- Becker T, Wenz LS, Kruger V, Lehmann W, Muller JM, Goroncy L, Zufall N, Lithgow T, Guiard B, Chacinska A, Wagner R, Meisinger C, Pfanner N. 2011. The mitochondrial import protein Mim1 promotes biogenesis of multispanning outer membrane proteins. J Cell Biol 194:387–395. doi:10.1083/jcb.201102044. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases