

Change of function of the wheat stress-responsive transcriptional repressor TaRAP2.1L by repressor motif modification
- PMID: 26150199
- PMCID: PMC11629789
- DOI: 10.1111/pbi.12432
Change of function of the wheat stress-responsive transcriptional repressor TaRAP2.1L by repressor motif modification
Abstract
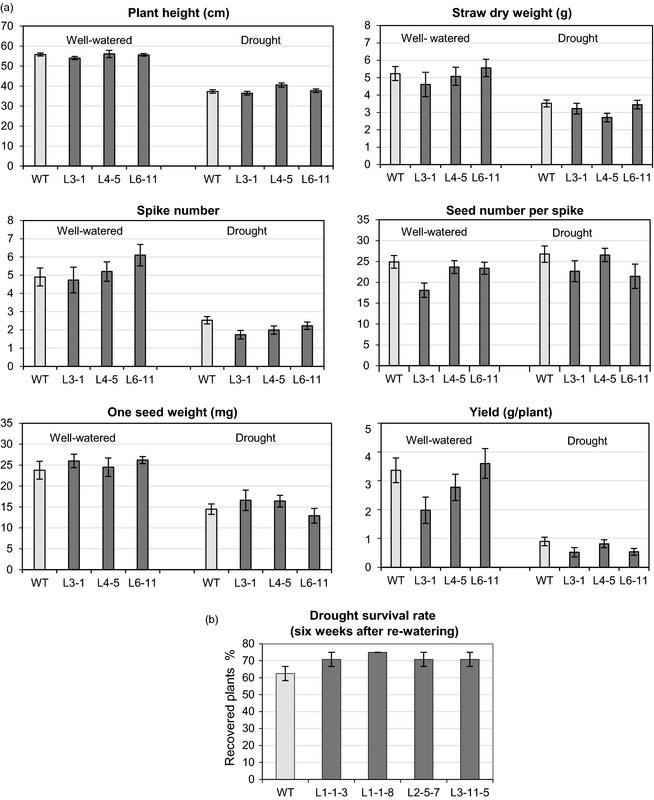
Plants respond to abiotic stresses by changes in gene regulation, including stress-inducible expression of transcriptional activators and repressors. One of the best characterized families of drought-related transcription factors are dehydration-responsive element binding (DREB) proteins, known as C-repeat binding factors (CBF). The wheat DREB/CBF gene TaRAP2.1L was isolated from drought-affected tissues using a dehydration-responsive element (DRE) as bait in a yeast one-hybrid screen. TaRAP2.1L is induced by elevated abscisic acid, drought and cold. A C-terminal ethylene responsive factor-associated amphiphilic repression (EAR) motif, known to be responsible for active repression of target genes, was identified in the TaRAP2.1L protein. It was found that TaRAP2.1L has a unique selectivity of DNA-binding, which differs from that of DREB activators. This binding selectivity remains unchanged in a TaRAP2.1L variant with an inactivated EAR motif (TaRAP2.1Lmut). To study the role of the TaRAP2.1L repressor activity associated with the EAR motif in planta, transgenic wheat overexpressing native or mutated TaRAP2.1L was generated. Overexpression of TaRAP2.1L under constitutive and stress-inducible promoters in transgenic wheat and barley led to dwarfism and decreased frost tolerance. By contrast, constitutive overexpression of the TaRAP2.1Lmut gene had little or no negative influence on wheat development or grain yield. Transgenic lines with the TaRAP2.1Lmut transgene had an enhanced ability to survive frost and drought. The improved stress tolerance is attributed to up-regulation of several stress-related genes known to be downstream genes of DREB/CBF activators.
Keywords: EAR motif; dehydration-responsive element binding proteins; drought; frost; transcriptional repressor; transgenic wheat.
© 2015 Society for Experimental Biology, Association of Applied Biologists and John Wiley & Sons Ltd.
Conflict of interest statement
The authors of this manuscript have no conflict of interest.
Figures







References
-
- Agarwal, P.K. , Agarwal, P. , Reddy, M.K. and Sopory, S.K. (2006) Role of DREB transcription factors in abiotic and biotic stress tolerance in plants. Plant Cell Rep. 25, 1263–1274. - PubMed
-
- Akhtar, M. , Jaiswal, A. , Taj, G. , Jaiswal, J.P. , Qureshi, M.I. and Singh, N.K. (2012) DREB1/CBF transcription factors: their structure, function and role in abiotic stress tolerance in plants. J. Genet. 91, 385–395. - PubMed
-
- Canella, D. , Gilmour, S.J. , Kuhn, L.A. and Thomashow, M.F. (2010) DNA‐binding by the Arabidopsis CBF1 transcription factor requires the PKKP/RAGRxKFxETRHP signature sequence. Biochim. Biophys. Acta, 1799, 454–462. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
