

Targeting Neuronal Networks with Combined Drug and Stimulation Paradigms Guided by Neuroimaging to Treat Brain Disorders
- PMID: 26150315
- PMCID: PMC6287502
- DOI: 10.1177/1073858415592377
Targeting Neuronal Networks with Combined Drug and Stimulation Paradigms Guided by Neuroimaging to Treat Brain Disorders
Abstract
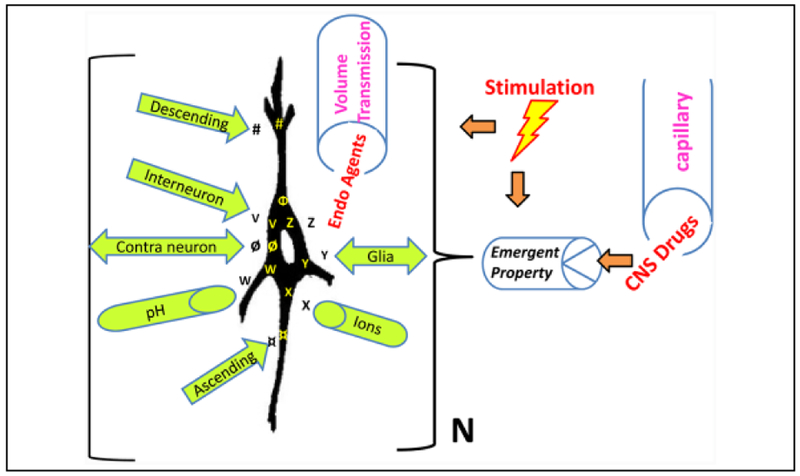
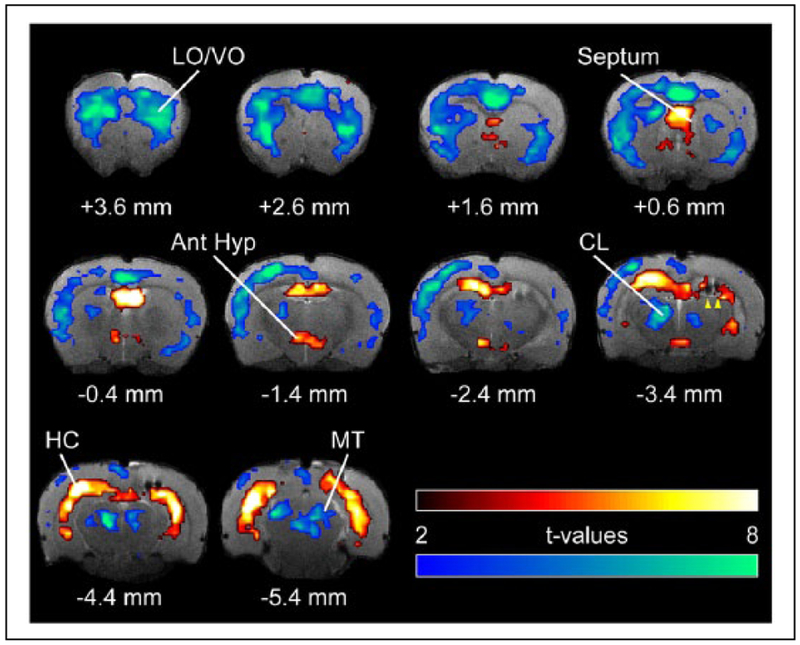
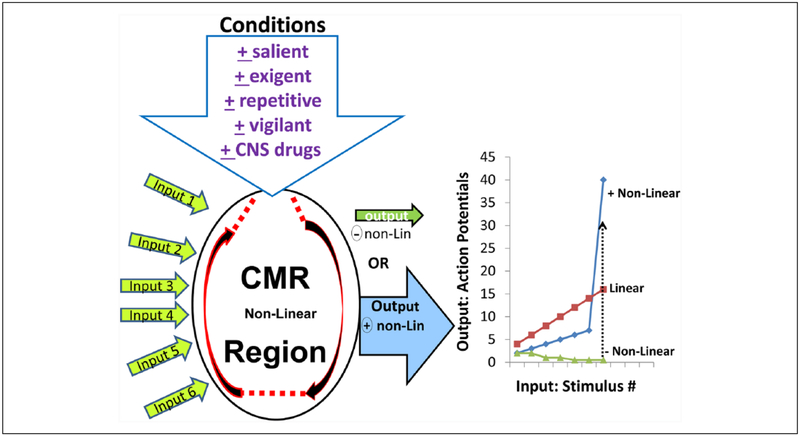
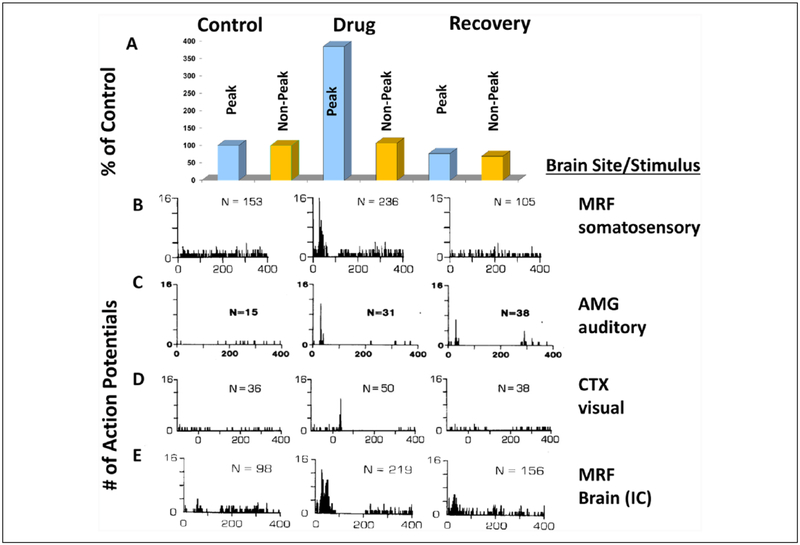
Improved therapy of brain disorders can be achieved by focusing on neuronal networks, utilizing combined pharmacological and stimulation paradigms guided by neuroimaging. Neuronal networks that mediate normal brain functions, such as hearing, interact with other networks, which is important but commonly neglected. Network interaction changes often underlie brain disorders, including epilepsy. "Conditional multireceptive" (CMR) brain areas (e.g., brainstem reticular formation and amygdala) are critical in mediating neuroplastic changes that facilitate network interactions. CMR neurons receive multiple inputs but exhibit extensive response variability due to milieu and behavioral state changes and are exquisitely sensitive to agents that increase or inhibit GABA-mediated inhibition. Enhanced CMR neuronal responsiveness leads to expression of emergent properties--nonlinear events--resulting from network self-organization. Determining brain disorder mechanisms requires animals that model behaviors and neuroanatomical substrates of human disorders identified by neuroimaging. However, not all sites activated during network operation are requisite for that operation. Other active sites are ancillary, because their blockade does not alter network function. Requisite network sites exhibit emergent properties that are critical targets for pharmacological and stimulation therapies. Improved treatment of brain disorders should involve combined pharmacological and stimulation therapies, guided by neuroimaging, to correct network malfunctions by targeting specific network neurons.
Keywords: CNS disorders; CNS pharmacology; complexity; electrical stimulation; emergent property; neuroimaging; neuronal network; pain; seizures; startle.
© The Author(s) 2015.
Conflict of interest statement
Declaration of Conflicting Interests
The authors declare no potential conflict of interest with respect to the research, authorship, and/or publication of this article.
Figures

Similar articles
-
Lethal Interactions of neuronal networks in epilepsy mediated by both synaptic and volume transmission indicate approaches to prevention.Prog Neurobiol. 2025 Jun;249:102770. doi: 10.1016/j.pneurobio.2025.102770. Epub 2025 Apr 19. Prog Neurobiol. 2025. PMID: 40258456 Review.
-
Electrical stimulation therapies for CNS disorders and pain are mediated by competition between different neuronal networks in the brain.Med Hypotheses. 2008 Nov;71(5):668-81. doi: 10.1016/j.mehy.2008.06.030. Epub 2008 Aug 30. Med Hypotheses. 2008. PMID: 18762389 Free PMC article.
-
Emergent properties of CNS neuronal networks as targets for pharmacology: application to anticonvulsant drug action.Prog Neurobiol. 2004 Jan;72(1):55-85. doi: 10.1016/j.pneurobio.2003.11.003. Prog Neurobiol. 2004. PMID: 15019176 Review.
-
Functional neurosurgery. The modulation of neural and mind circuits.Neurosciences (Riyadh). 2012 Jan;17(1):16-31. Neurosciences (Riyadh). 2012. PMID: 22246006 Review.
-
Neuronal networks in the genetically epilepsy-prone rat.Adv Neurol. 1999;79:311-21. Adv Neurol. 1999. PMID: 10514823 Review.
Cited by
-
Lethal Interactions of neuronal networks in epilepsy mediated by both synaptic and volume transmission indicate approaches to prevention.Prog Neurobiol. 2025 Jun;249:102770. doi: 10.1016/j.pneurobio.2025.102770. Epub 2025 Apr 19. Prog Neurobiol. 2025. PMID: 40258456 Review.
-
Bypassing the Blood-Brain Barrier: Direct Intracranial Drug Delivery in Epilepsies.Pharmaceutics. 2020 Nov 24;12(12):1134. doi: 10.3390/pharmaceutics12121134. Pharmaceutics. 2020. PMID: 33255396 Free PMC article. Review.
-
Vascular Cognitive Impairment After Mild Stroke: Connectomic Insights, Neuroimaging, and Knowledge Translation.Front Neurosci. 2022 Jul 7;16:905979. doi: 10.3389/fnins.2022.905979. eCollection 2022. Front Neurosci. 2022. PMID: 35937885 Free PMC article. Review.
-
Optogenetic study of networks in epilepsy.J Neurosci Res. 2017 Dec;95(12):2325-2335. doi: 10.1002/jnr.23767. Epub 2016 Jul 14. J Neurosci Res. 2017. PMID: 27413006 Free PMC article. Review.
-
Cognitive impairment after traumatic brain injury is associated with reduced long-term depression of excitatory postsynaptic potential in the rat hippocampal dentate gyrus.Neural Regen Res. 2018 Oct;13(10):1753-1758. doi: 10.4103/1673-5374.238618. Neural Regen Res. 2018. PMID: 30136690 Free PMC article.
References
-
- Agnati LF, Genedani S, Spano P, Guidolin D, Fuxe K. 2014. Volume transmission and the Russian-doll organization of brain cell networks: aspects of their integrative actions In: Faingold C, Blumenfeld H, editors. Neuronal networks in brain function, CNS disorders, and therapeutics. San Diego, CA: Academic Press; p. 103–19.
-
- Allen L, Mehta S, McClure JA, Teasell R. 2012. Therapeutic interventions for aphasia initiated more than six months post stroke: a review of the evidence. Top Stroke Rehabil 19(6):523–35. - PubMed
-
- Barbarisi M, Pace MC, Passavanti MB, Maisto M, Mazzariello L, Pota V, and others. 2010. Pregabalin and transcutaneous electrical nerve stimulation for postherpetic neuralgia treatment. Clin J Pain 26(7):567–72. - PubMed
-
- Bellani M, Dusi N, Yeh PH, Soares JC, Brambilla P. 2011. The effects of antidepressants on human brain as detected by imaging studies. Focus on major depression. Prog Neuropsychopharmacol Biol Psychiatry 35(7):1544–52. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
