

Interphase centrosome organization by the PLP-Cnn scaffold is required for centrosome function
- PMID: 26150390
- PMCID: PMC4494003
- DOI: 10.1083/jcb.201503117
Interphase centrosome organization by the PLP-Cnn scaffold is required for centrosome function
Abstract
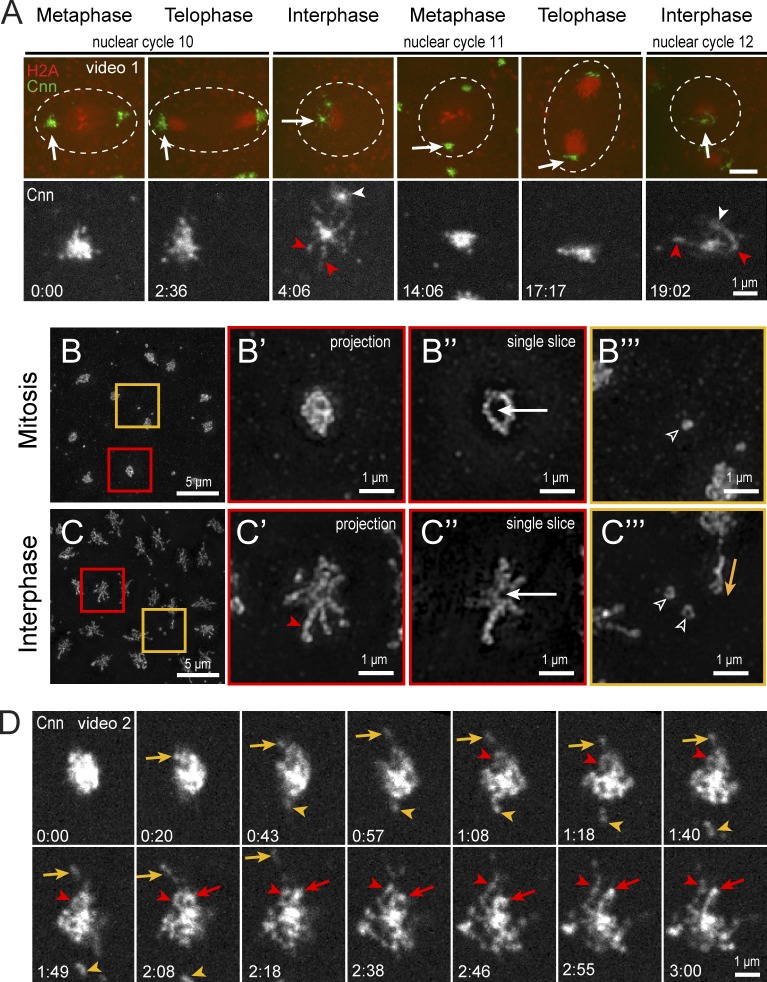
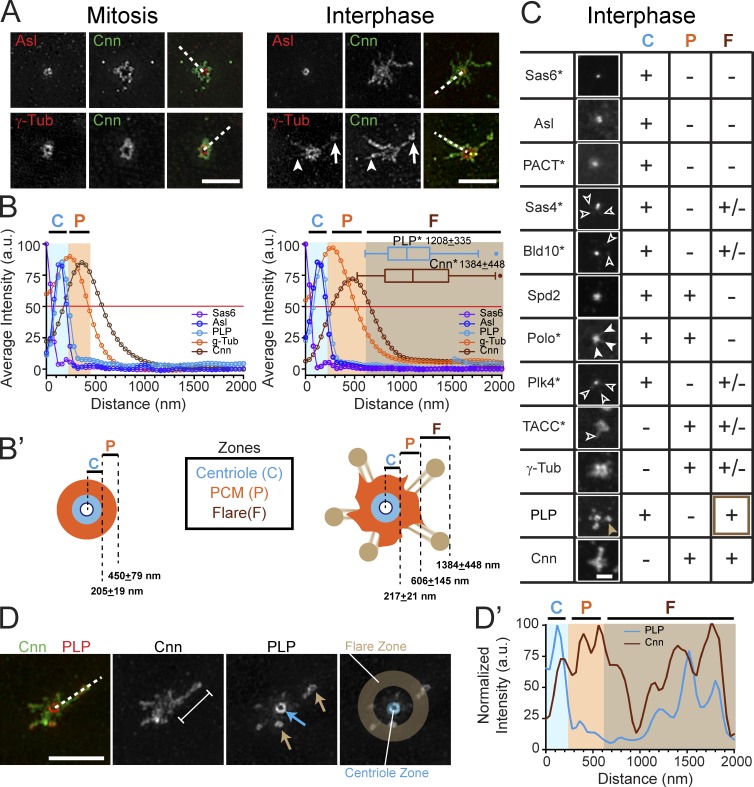
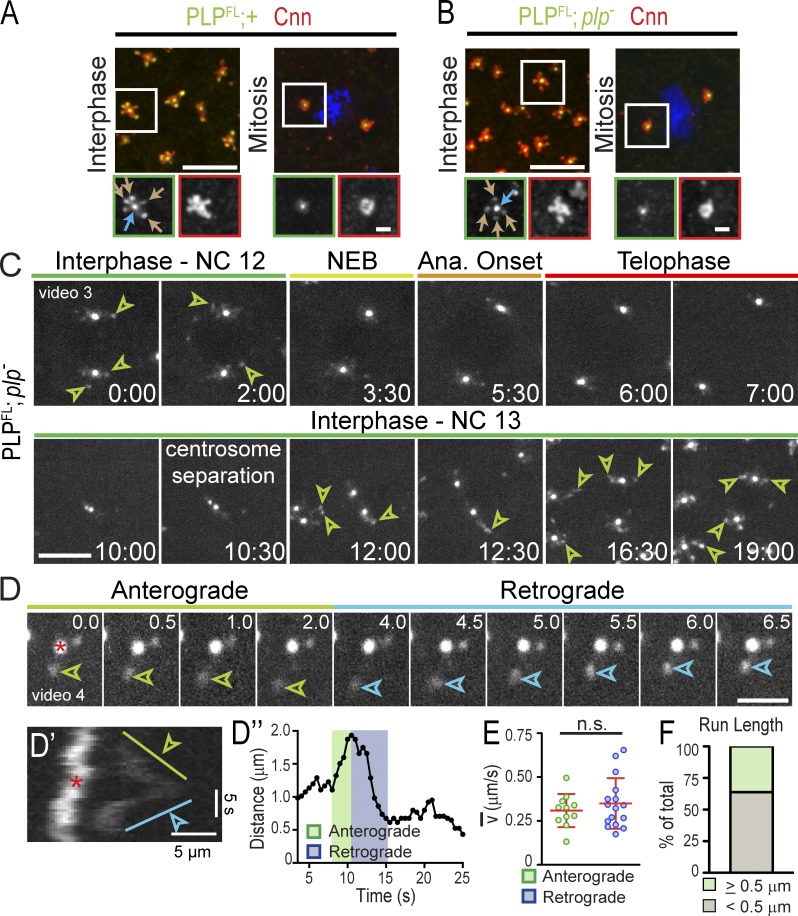
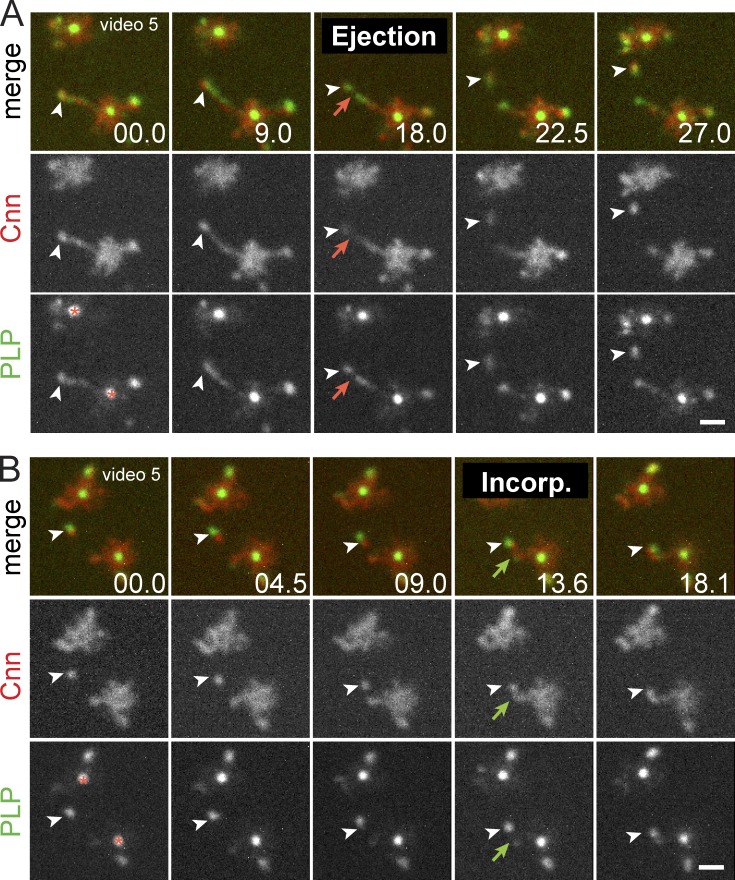
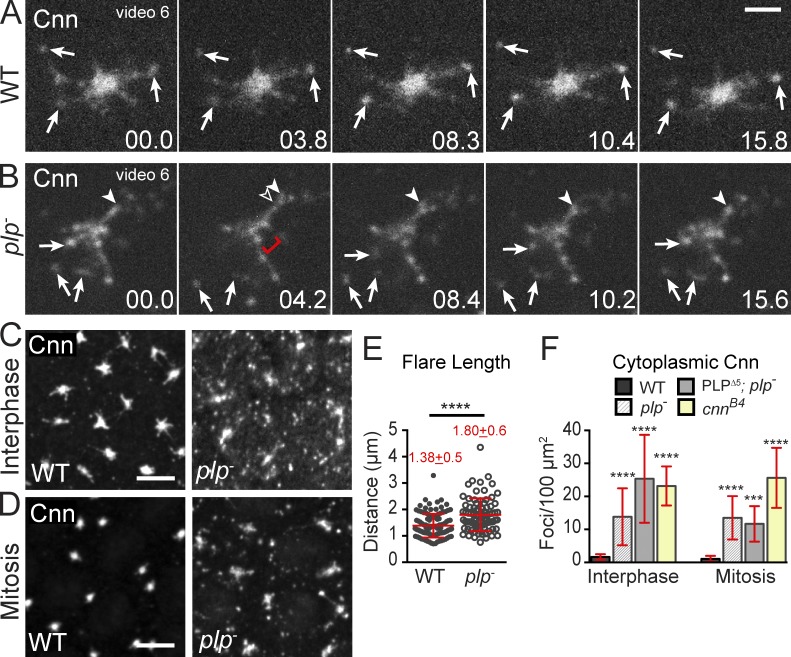
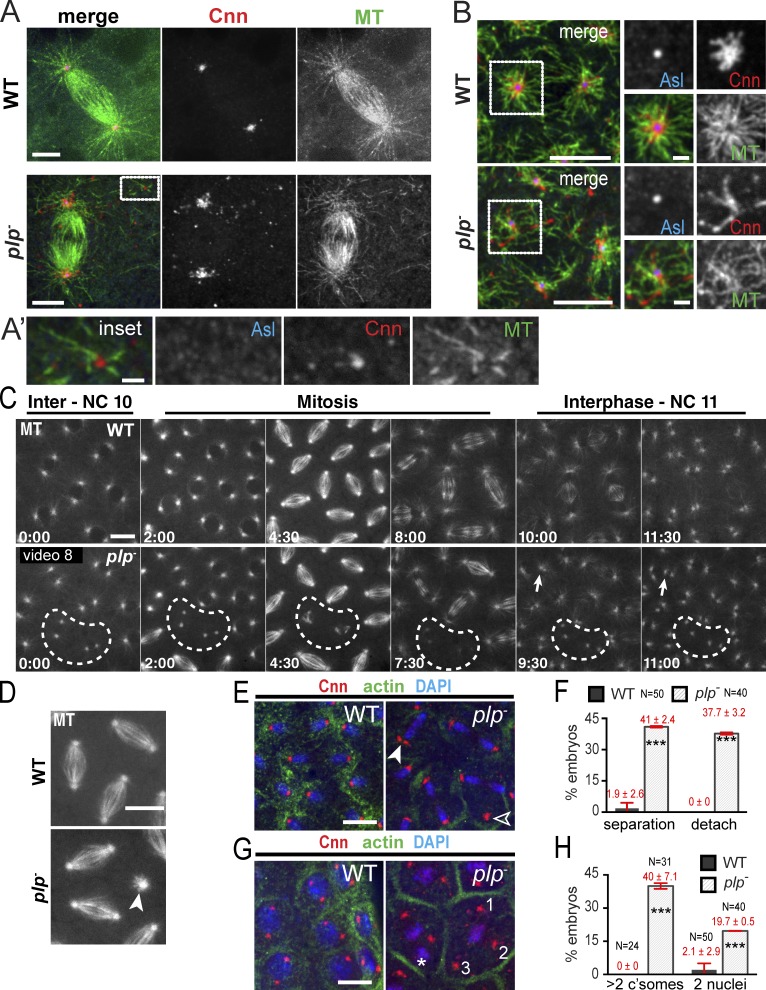
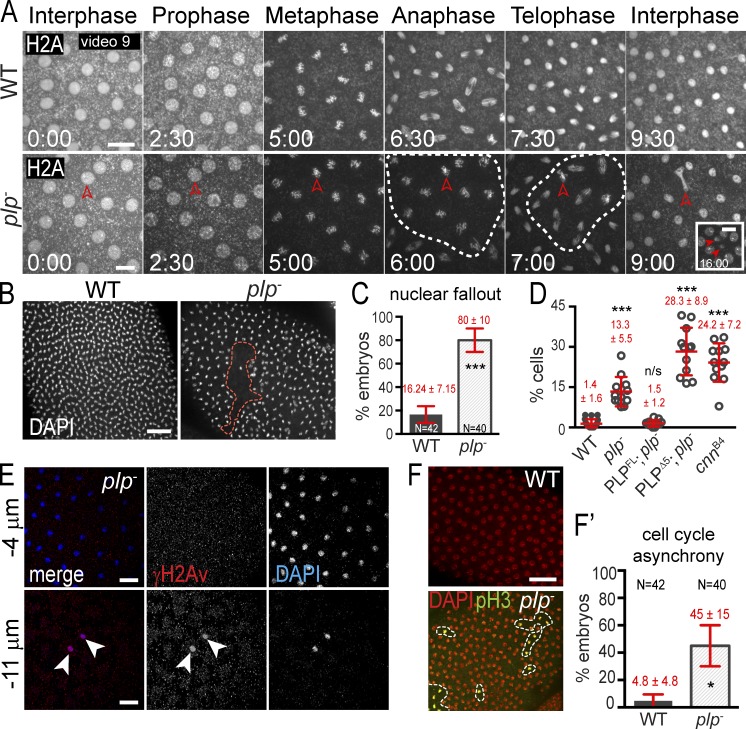
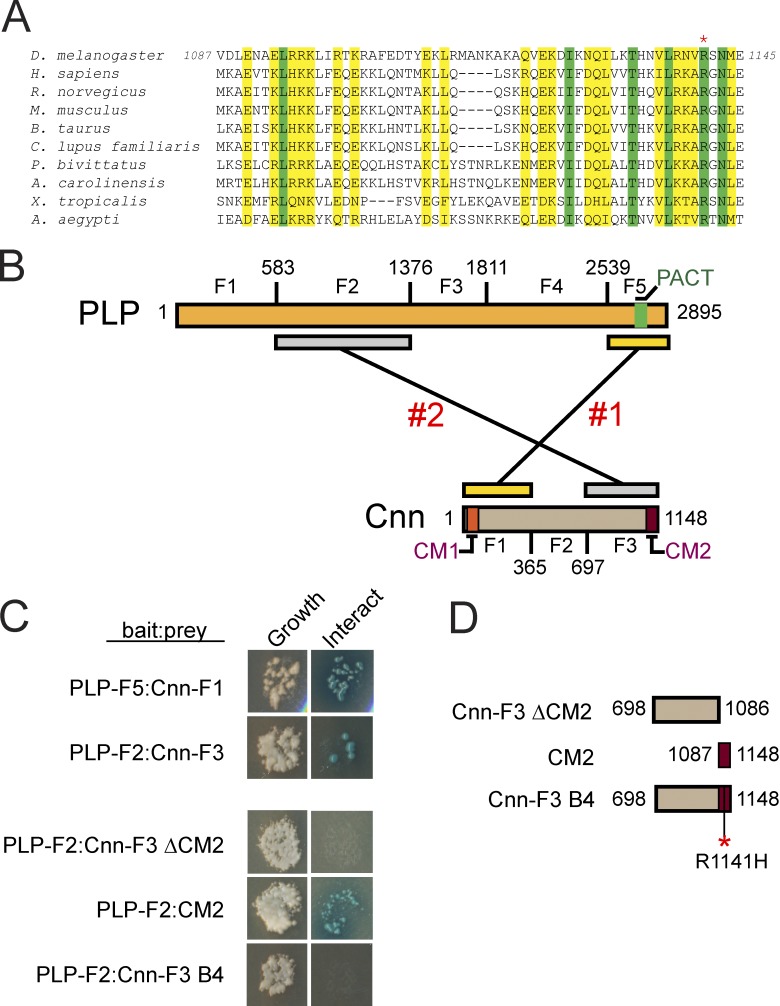
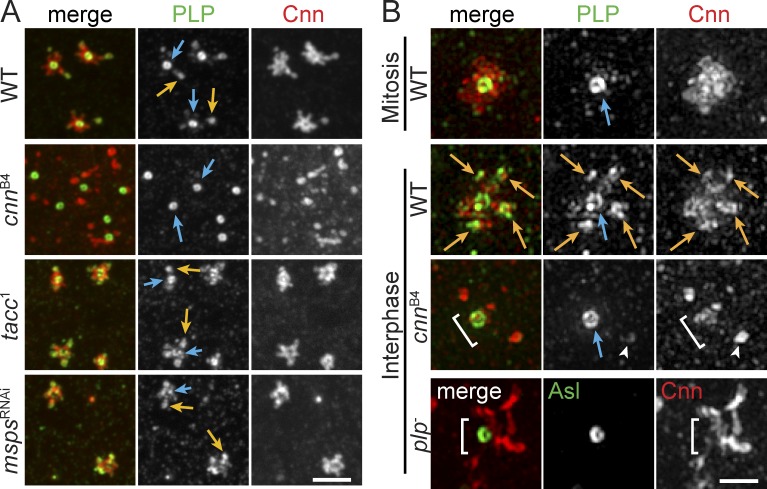
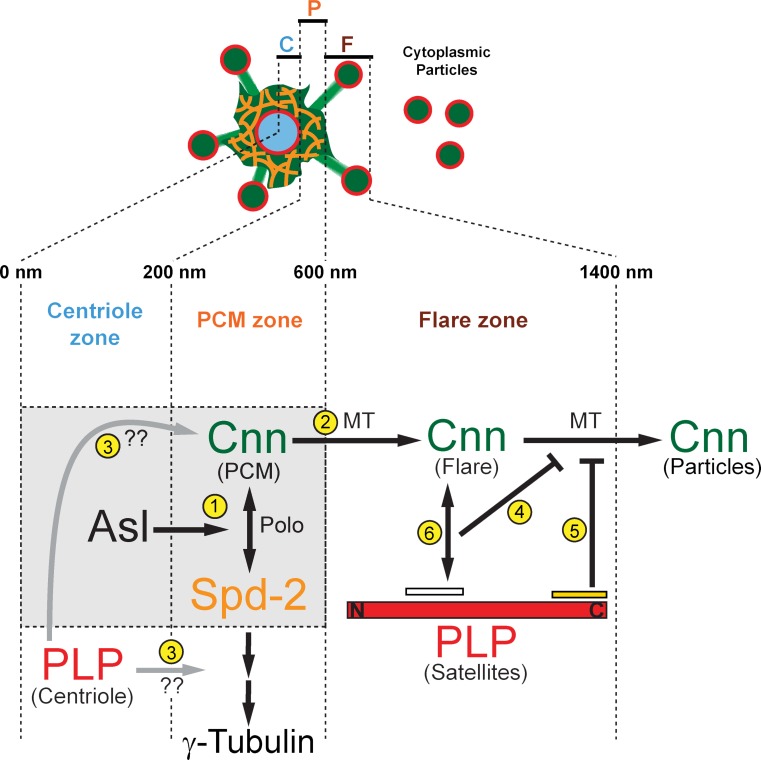
Pericentriolar material (PCM) mediates the microtubule (MT) nucleation and anchoring activity of centrosomes. A scaffold organized by Centrosomin (Cnn) serves to ensure proper PCM architecture and functional changes in centrosome activity with each cell cycle. Here, we investigate the mechanisms that spatially restrict and temporally coordinate centrosome scaffold formation. Focusing on the mitotic-to-interphase transition in Drosophila melanogaster embryos, we show that the elaboration of the interphase Cnn scaffold defines a major structural rearrangement of the centrosome. We identify an unprecedented role for Pericentrin-like protein (PLP), which localizes to the tips of extended Cnn flares, to maintain robust interphase centrosome activity and promote the formation of interphase MT asters required for normal nuclear spacing, centrosome segregation, and compartmentalization of the syncytial embryo. Our data reveal that Cnn and PLP directly interact at two defined sites to coordinate the cell cycle-dependent rearrangement and scaffolding activity of the centrosome to permit normal centrosome organization, cell division, and embryonic viability.
Figures
References
-
- Bond J., Roberts E., Springell K., Lizarraga S.B., Scott S., Higgins J., Hampshire D.J., Morrison E.E., Leal G.F., Silva E.O., et al. . 2005. A centrosomal mechanism involving CDK5RAP2 and CENPJ controls brain size. Nat. Genet. 37:353–355. (published erratum appears in Nat. Genet. 2005. 37:555) 10.1038/ng1539 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
