

Schwann cell autophagy, myelinophagy, initiates myelin clearance from injured nerves
- PMID: 26150392
- PMCID: PMC4494002
- DOI: 10.1083/jcb.201503019
Schwann cell autophagy, myelinophagy, initiates myelin clearance from injured nerves
Abstract
Although Schwann cell myelin breakdown is the universal outcome of a remarkably wide range of conditions that cause disease or injury to peripheral nerves, the cellular and molecular mechanisms that make Schwann cell-mediated myelin digestion possible have not been established. We report that Schwann cells degrade myelin after injury by a novel form of selective autophagy, myelinophagy. Autophagy was up-regulated by myelinating Schwann cells after nerve injury, myelin debris was present in autophagosomes, and pharmacological and genetic inhibition of autophagy impaired myelin clearance. Myelinophagy was positively regulated by the Schwann cell JNK/c-Jun pathway, a central regulator of the Schwann cell reprogramming induced by nerve injury. We also present evidence that myelinophagy is defective in the injured central nervous system. These results reveal an important role for inductive autophagy during Wallerian degeneration, and point to potential mechanistic targets for accelerating myelin clearance and improving demyelinating disease.
© 2015 Gomez-Sanchez et al.
Figures
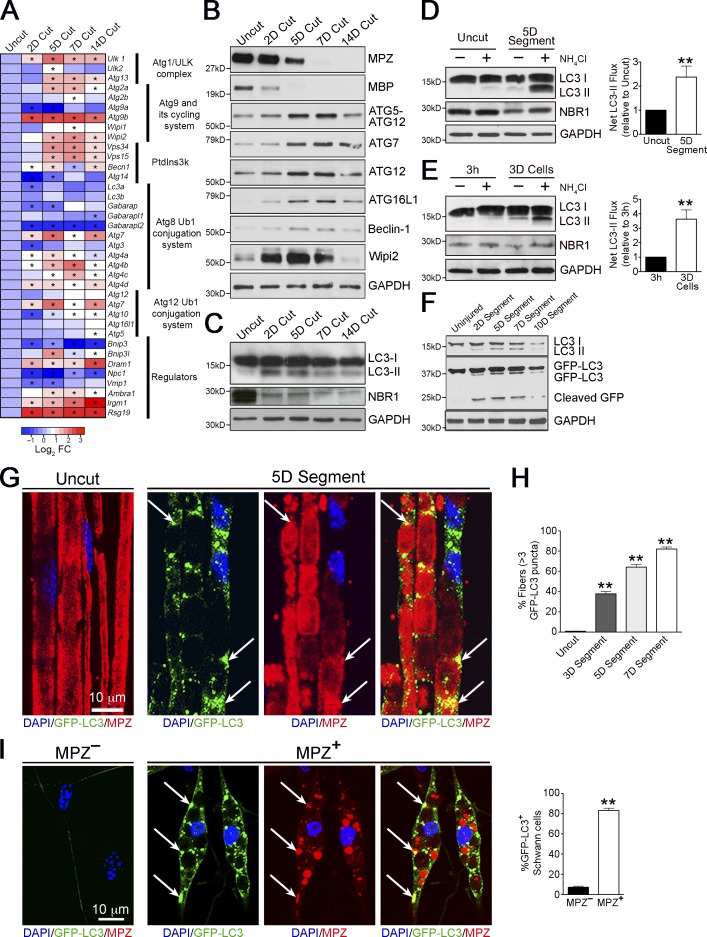
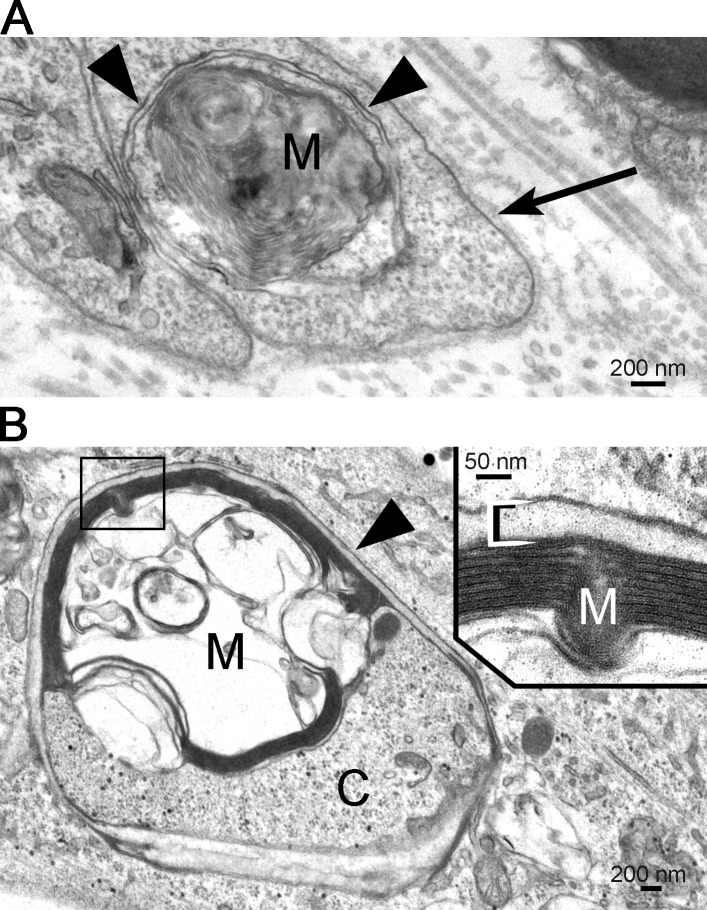
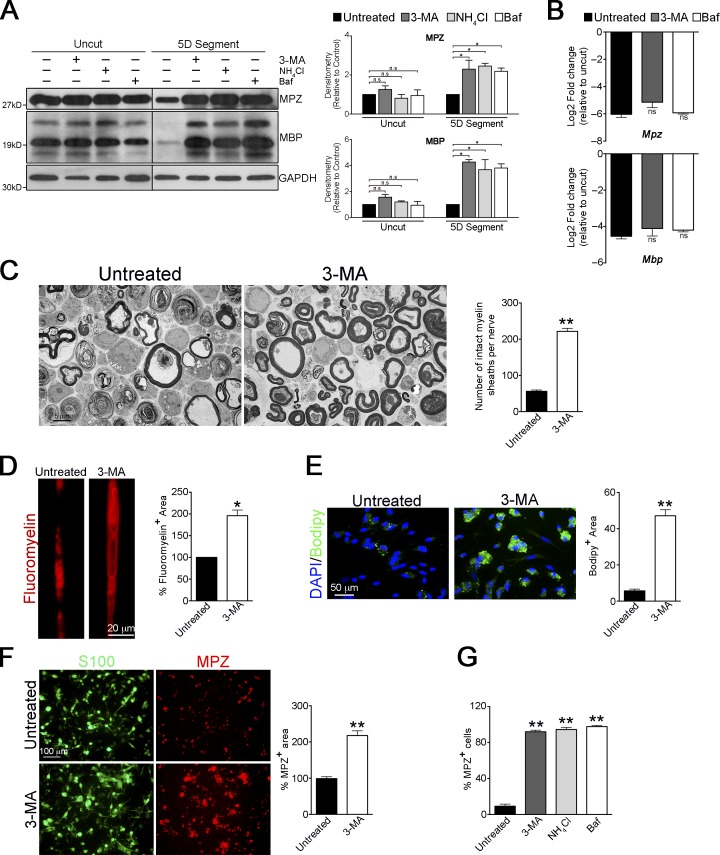
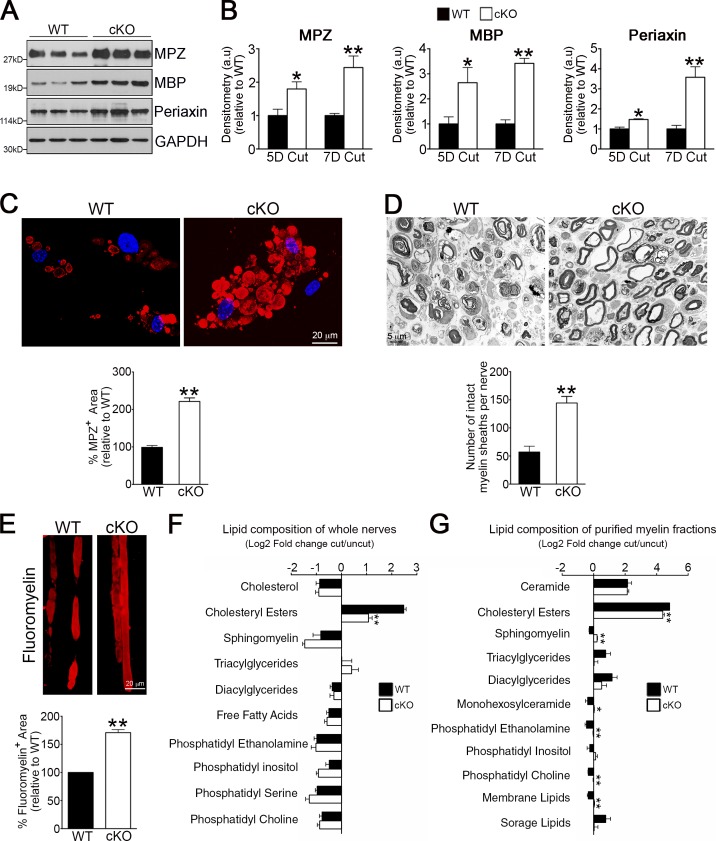
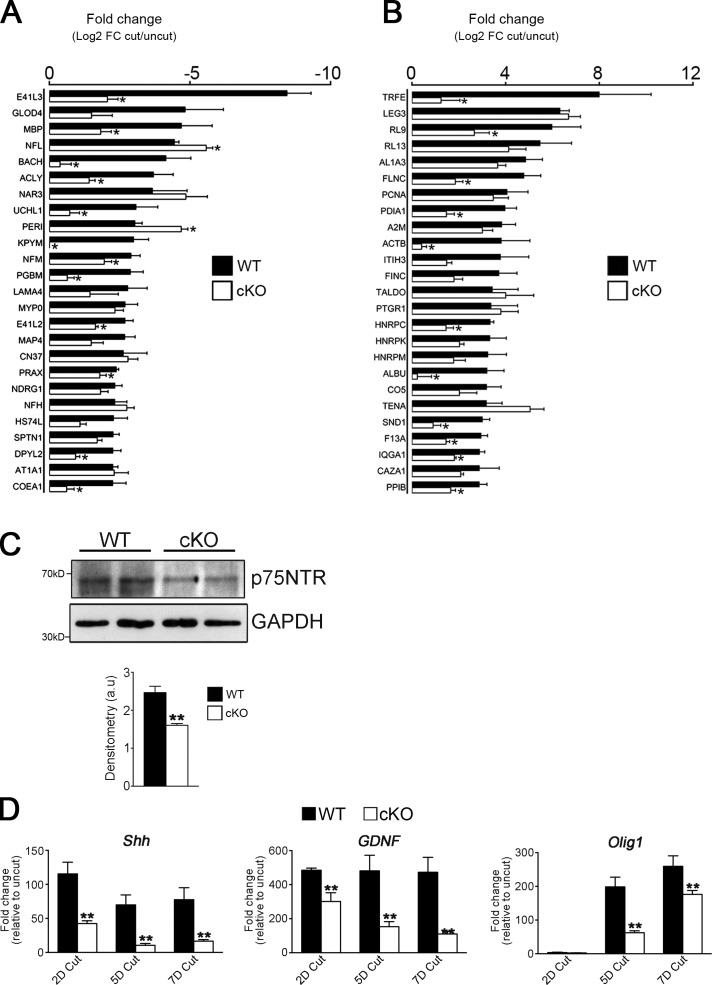
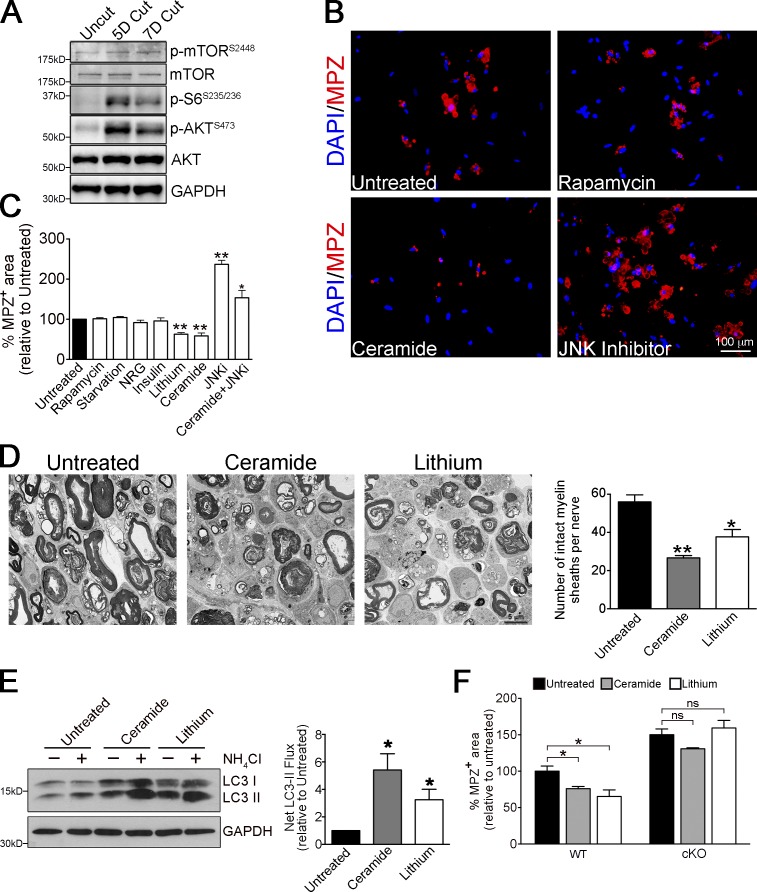
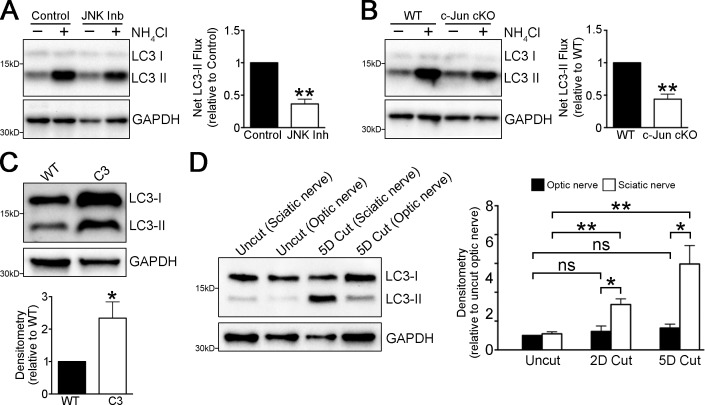
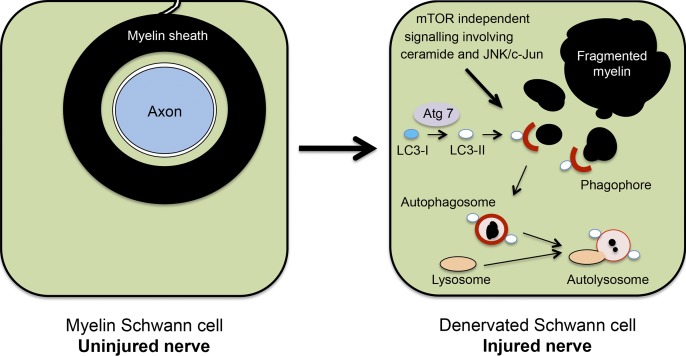
Comment in
-
Myelinophagy: Schwann cells dine in.J Cell Biol. 2015 Jul 6;210(1):9-10. doi: 10.1083/jcb.201506039. J Cell Biol. 2015. PMID: 26150387 Free PMC article.
References
-
- Arthur-Farraj P.J., Latouche M., Wilton D.K., Quintes S., Chabrol E., Banerjee A., Woodhoo A., Jenkins B., Rahman M., Turmaine M., et al. . 2012. c-Jun reprograms Schwann cells of injured nerves to generate a repair cell essential for regeneration. Neuron. 75:633–647. 10.1016/j.neuron.2012.06.021 - DOI - PMC - PubMed
-
- Barr J., Caballería J., Martínez-Arranz I., Domínguez-Díez A., Alonso C., Muntané J., Pérez-Cormenzana M., García-Monzón C., Mayo R., Martín-Duce A., et al. . 2012. Obesity-dependent metabolic signatures associated with nonalcoholic fatty liver disease progression. J. Proteome Res. 11:2521–2532. 10.1021/pr201223p - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
