

Structural basis of the mercury(II)-mediated conformational switching of the dual-function transcriptional regulator MerR
- PMID: 26150423
- PMCID: PMC4551924
- DOI: 10.1093/nar/gkv681
Structural basis of the mercury(II)-mediated conformational switching of the dual-function transcriptional regulator MerR
Abstract
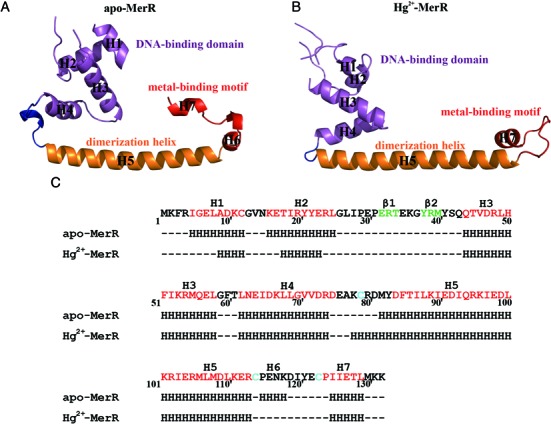
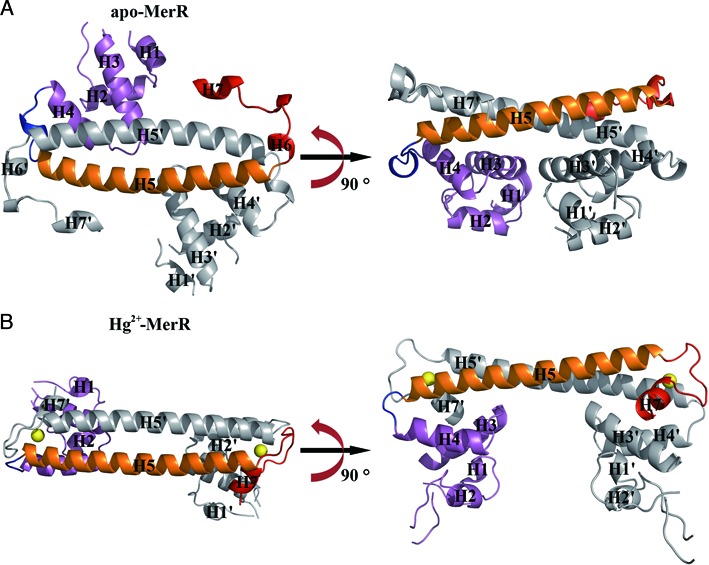
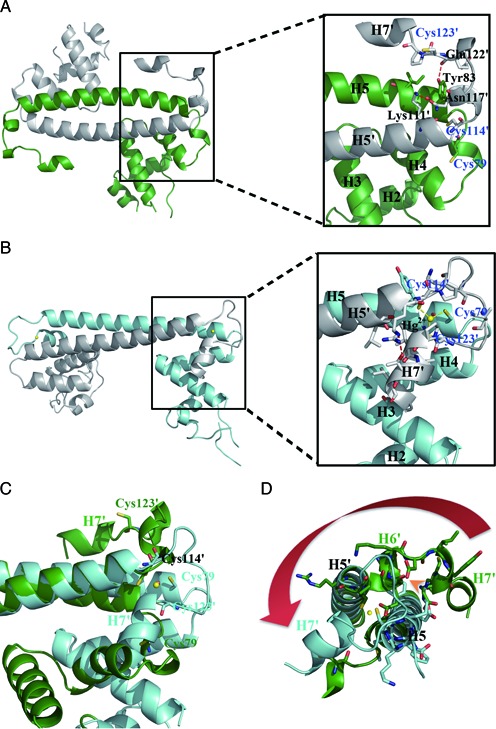
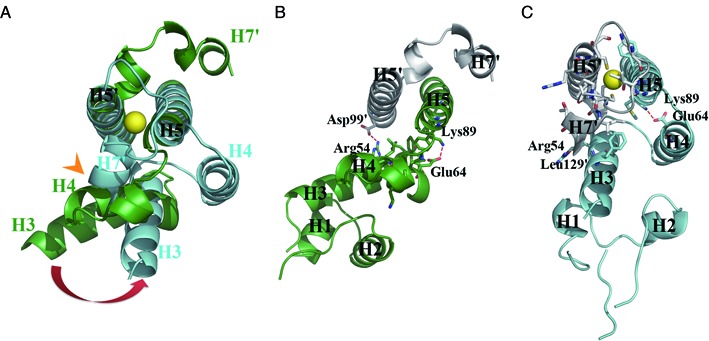
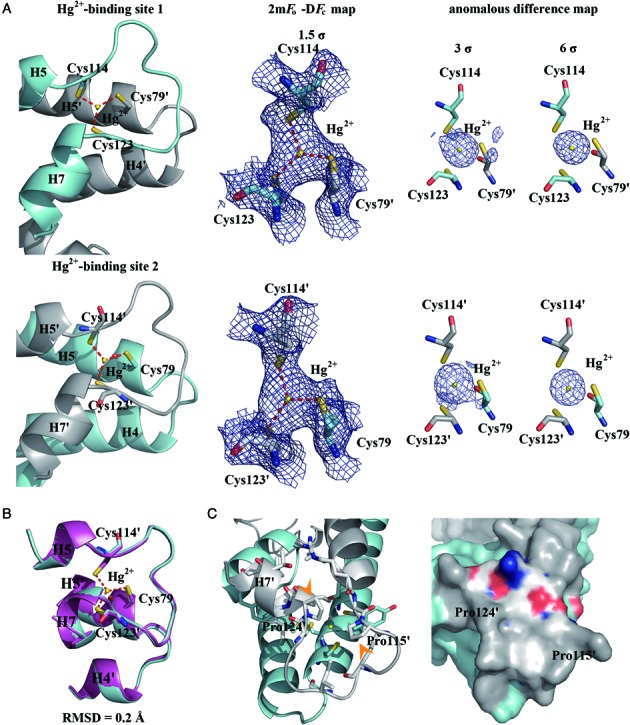
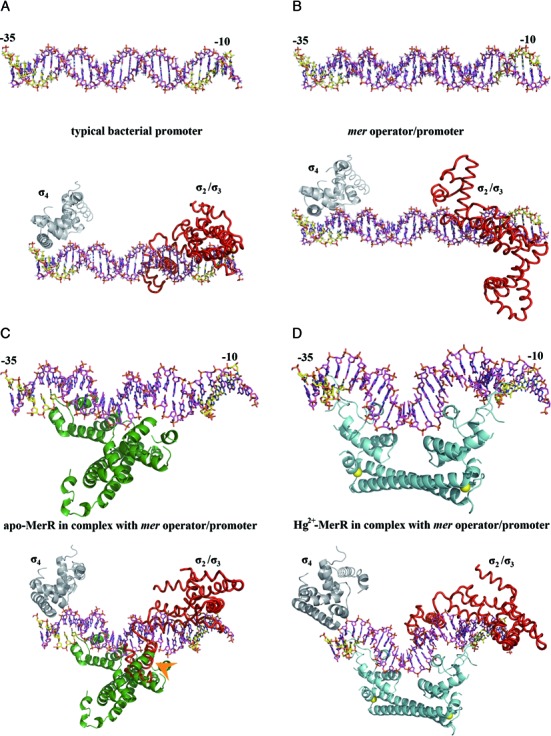
The mer operon confers bacterial resistance to inorganic mercury (Hg(2+)) and organomercurials by encoding proteins involved in sensing, transport and detoxification of these cytotoxic agents. Expression of the mer operon is under tight control by the dual-function transcriptional regulator MerR. The metal-free, apo MerR binds to the mer operator/promoter region as a repressor to block transcription initiation, but is converted into an activator upon Hg(2+)-binding. To understand how MerR interacts with Hg(2+) and how Hg(2+)-binding modulates MerR function, we report here the crystal structures of apo and Hg(2+)-bound MerR from Bacillus megaterium, corresponding respectively to the repressor and activator conformation of MerR. To our knowledge, the apo-MerR structure represents the first visualization of a MerR family member in its intact and inducer-free form. And the Hg(2+)-MerR structure offers the first view of a triligated Hg(2+)-thiolate center in a metalloprotein, confirming that MerR binds Hg(2+) via trigonal planar coordination geometry. Structural comparison revealed the conformational transition of MerR is coupled to the assembly/disassembly of a buried Hg(2+) binding site, thereby providing a structural basis for the Hg(2+)-mediated functional switching of MerR. The pronounced Hg(2+)-induced repositioning of the MerR DNA-binding domains suggests a plausible mechanism for the transcriptional regulation of the mer operon.
© The Author(s) 2015. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Bacterial resistances to inorganic mercury salts and organomercurials.Plasmid. 1992 Jan;27(1):4-16. doi: 10.1016/0147-619x(92)90002-r. Plasmid. 1992. PMID: 1311113 Review.
-
In vivo DNA-protein interactions at the divergent mercury resistance (mer) promoters. II. Repressor/activator (MerR)-RNA polymerase interaction with merOP mutants.J Biol Chem. 1993 Feb 5;268(4):2632-9. J Biol Chem. 1993. PMID: 8428940
-
In vivo DNA-protein interactions at the divergent mercury resistance (mer) promoters. I. Metalloregulatory protein MerR mutants.J Biol Chem. 1993 Feb 5;268(4):2623-31. J Biol Chem. 1993. PMID: 8428939
-
Cd(II)-responsive and constitutive mutants implicate a novel domain in MerR.J Bacteriol. 1999 Jun;181(11):3462-71. doi: 10.1128/JB.181.11.3462-3471.1999. J Bacteriol. 1999. PMID: 10348859 Free PMC article.
-
Structural determinants of metal selectivity in prokaryotic metal-responsive transcriptional regulators.Biometals. 2005 Aug;18(4):413-28. doi: 10.1007/s10534-005-3716-8. Biometals. 2005. PMID: 16158234 Review.
Cited by
-
Molecular Evolution of Extensively Drug-Resistant (XDR) Pseudomonas aeruginosa Strains From Patients and Hospital Environment in a Prolonged Outbreak.Front Microbiol. 2019 Aug 8;10:1742. doi: 10.3389/fmicb.2019.01742. eCollection 2019. Front Microbiol. 2019. PMID: 31440214 Free PMC article.
-
AsIII Selectively Induces a Disorder-to-Order Transition in the Metalloid Binding Region of the AfArsR Protein.J Am Chem Soc. 2024 Jun 26;146(25):17009-17022. doi: 10.1021/jacs.3c11665. Epub 2024 May 31. J Am Chem Soc. 2024. PMID: 38820242 Free PMC article.
-
A Crystallographic Examination of Predisposition versus Preorganization in de Novo Designed Metalloproteins.J Am Chem Soc. 2016 Sep 14;138(36):11979-88. doi: 10.1021/jacs.6b07165. Epub 2016 Sep 2. J Am Chem Soc. 2016. PMID: 27532255 Free PMC article.
-
New Insights into Alterations in PL Proteins Affecting Their Binding to DNA after Exposure of Mytilus galloprovincialis to Mercury-A Possible Risk to Sperm Chromatin Structure?Int J Mol Sci. 2021 May 31;22(11):5893. doi: 10.3390/ijms22115893. Int J Mol Sci. 2021. PMID: 34072703 Free PMC article.
-
A New Facet of Vitamin B12: Gene Regulation by Cobalamin-Based Photoreceptors.Annu Rev Biochem. 2017 Jun 20;86:485-514. doi: 10.1146/annurev-biochem-061516-044500. Annu Rev Biochem. 2017. PMID: 28654327 Free PMC article. Review.
References
-
- Silver S., Phung le T. A bacterial view of the periodic table: genes and proteins for toxic inorganic ions. J. Ind. Microbiol. Biotechnol. 2005;32:587–605. - PubMed
-
- Giedroc D.P., Arunkumar A.I. Metal sensor proteins: nature's metalloregulated allosteric switches. Dalton Trans. 2007;29:3107–3120. - PubMed
-
- Barkay T., Miller S.M., Summers A.O. Bacterial mercury resistance from atoms to ecosystems. FEMS Microbiol. Rev. 2003;27:355–384. - PubMed
-
- Huang C.C., Narita M., Yamagata T., Itoh Y., Endo G. Structure analysis of a class II transposon encoding the mercury resistance of the Gram-positive Bacterium bacillus megaterium MB1, a strain isolated from minamata bay, Japan. Gene. 1999;234:361–369. - PubMed
-
- Brown N.L., Misra T.K., Winnie J.N., Schmidt A., Seiff M., Silver S. The nucleotide sequence of the mercuric resistance operons of plasmid R100 and transposon Tn501: further evidence for mer genes which enhance the activity of the mercuric ion detoxification system. Mol. Gen. Genet. 1986;202:143–151. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
