

The within-host dynamics of African trypanosome infections
- PMID: 26150654
- PMCID: PMC4528486
- DOI: 10.1098/rstb.2014.0288
The within-host dynamics of African trypanosome infections
Abstract
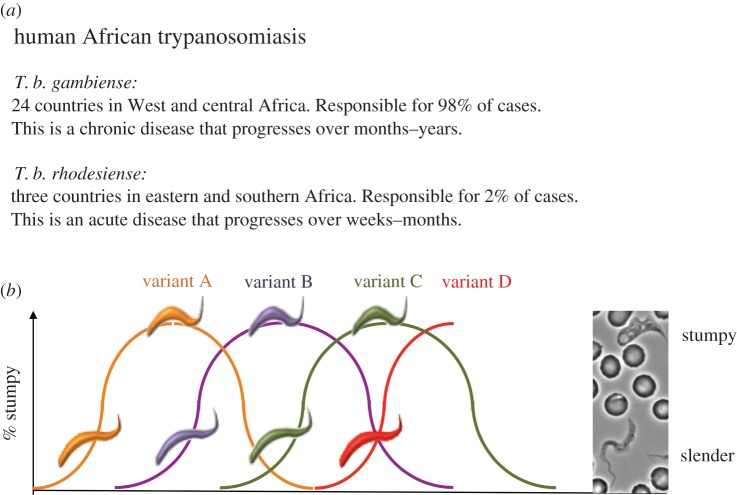
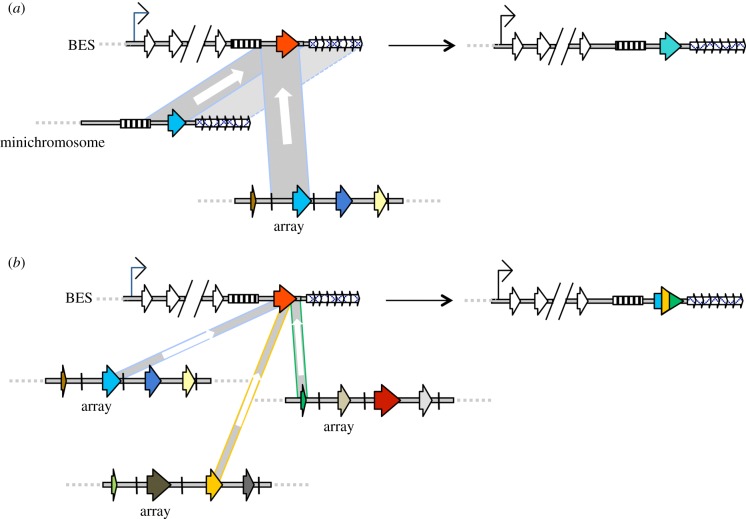
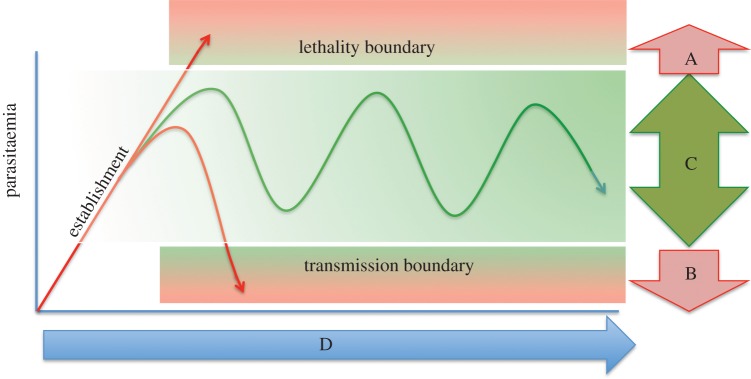
African trypanosomes are single-celled protozoan parasites that are capable of long-term survival while living extracellularly in the bloodstream and tissues of mammalian hosts. Prolonged infections are possible because trypanosomes undergo antigenic variation-the expression of a large repertoire of antigenically distinct surface coats, which allows the parasite population to evade antibody-mediated elimination. The mechanisms by which antigen genes become activated influence their order of expression, most likely by influencing the frequency of productive antigen switching, which in turn is likely to contribute to infection chronicity. Superimposed upon antigen switching as a contributor to trypanosome infection dynamics is the density-dependent production of cell-cycle arrested parasite transmission stages, which limit the infection while ensuring parasite spread to new hosts via the bite of blood-feeding tsetse flies. Neither antigen switching nor developmental progression to transmission stages is driven by the host. However, the host can contribute to the infection dynamic through the selection of distinct antigen types, the influence of genetic susceptibility or trypanotolerance and the potential influence of host-dependent effects on parasite virulence, development of transmission stages and pathogenicity. In a zoonotic infection cycle where trypanosomes circulate within a range of host animal populations, and in some cases humans, there is considerable scope for a complex interplay between parasite immune evasion, transmission potential and host factors to govern the profile and outcome of infection.
Keywords: Trypanosoma; antigenic variation; quorum sensing; transmission; trypanosome; within-host dynamics.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources