

The Mechanisms of Generation, Recognition, and Erasure of DNA 5-Methylcytosine and Thymine Oxidations
- PMID: 26152719
- PMCID: PMC4543634
- DOI: 10.1074/jbc.R115.656884
The Mechanisms of Generation, Recognition, and Erasure of DNA 5-Methylcytosine and Thymine Oxidations
Abstract
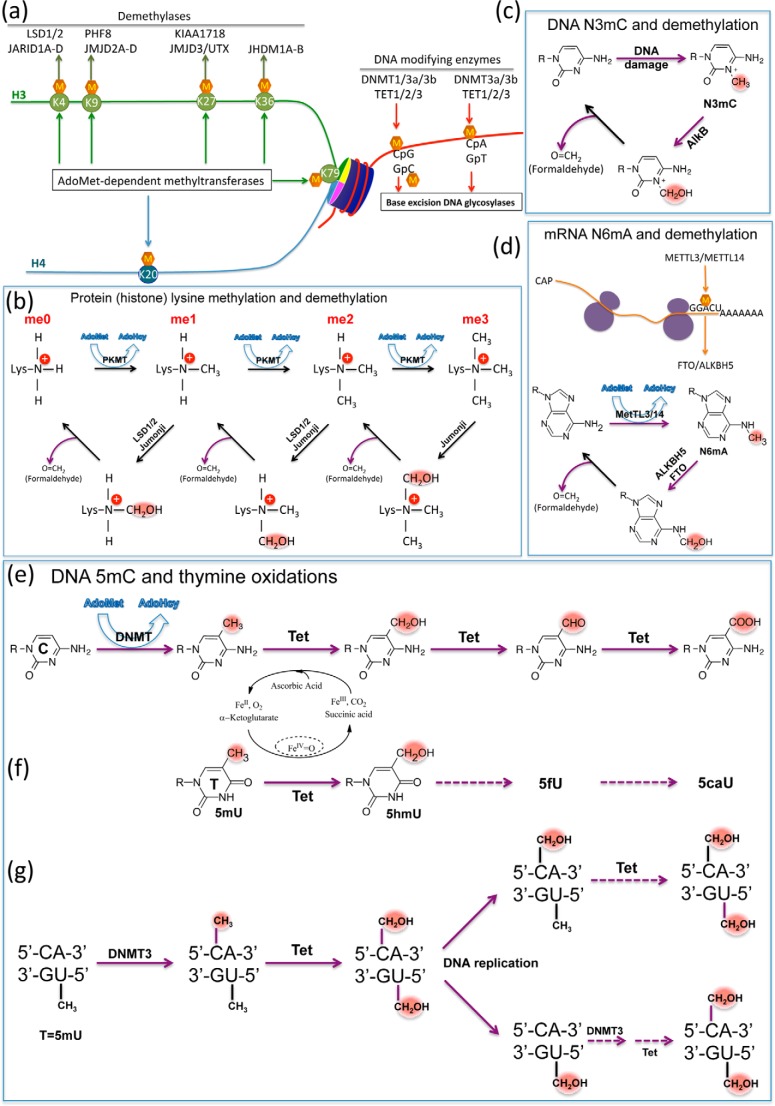
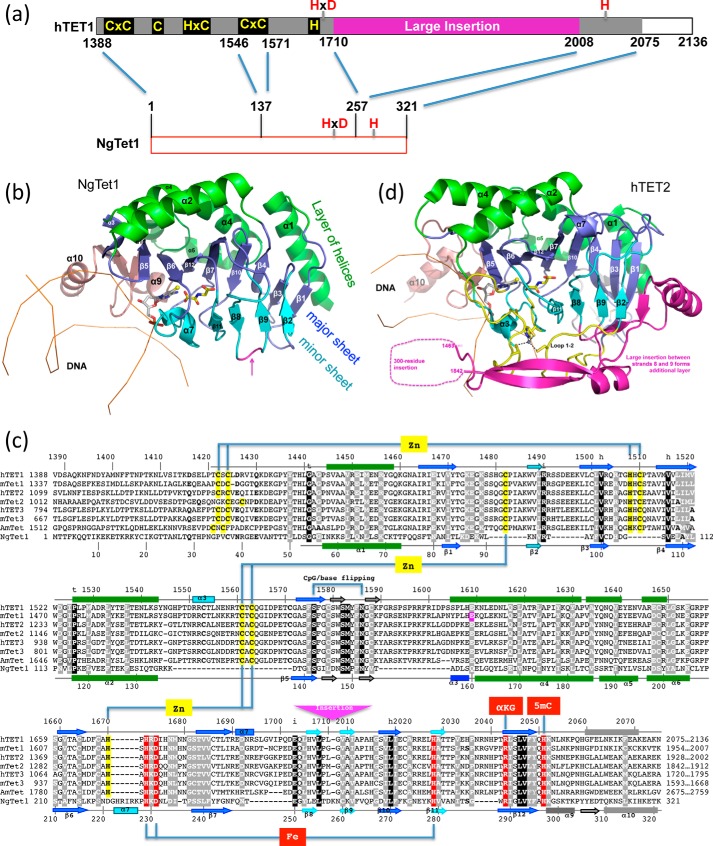
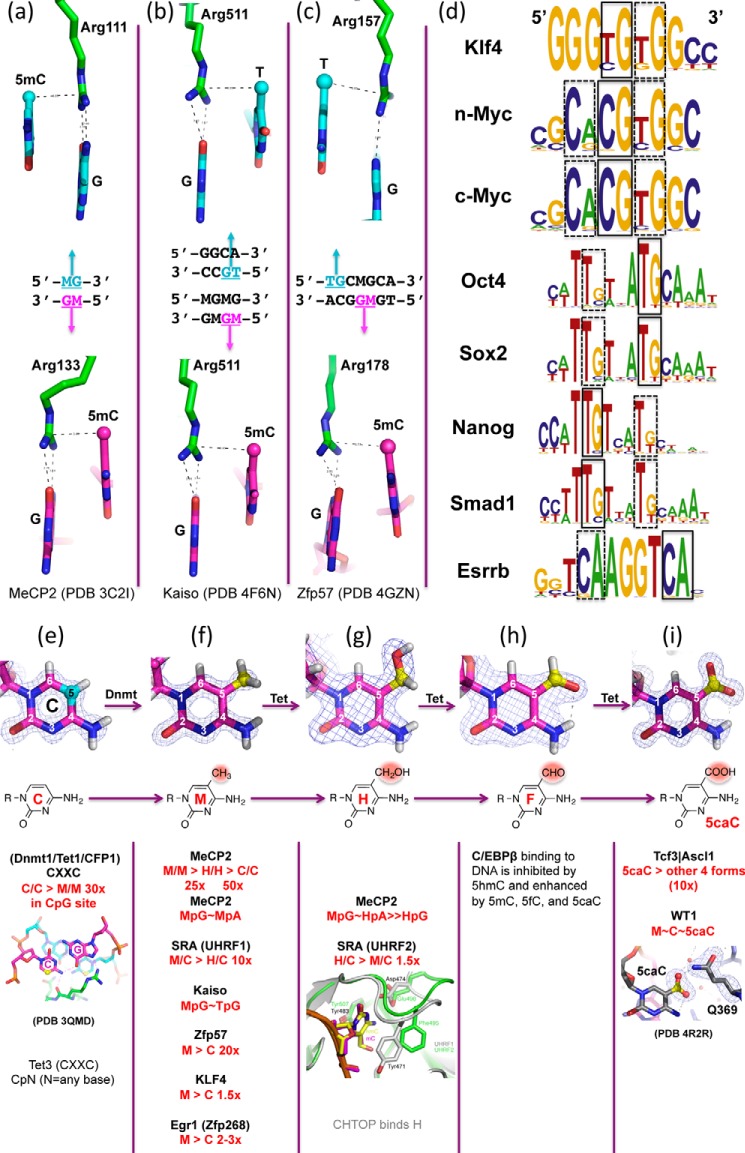
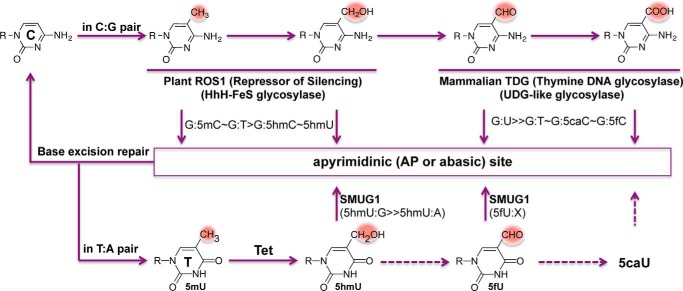
One of the most fundamental questions in the control of gene expression in mammals is how the patterns of epigenetic modifications of DNA are generated, recognized, and erased. This includes covalent cytosine methylation of DNA and its associated oxidation states. An array of AdoMet-dependent methyltransferases, Fe(II)- and α-ketoglutarate-dependent dioxygenases, base excision glycosylases, and sequence-specific transcription factors is responsible for changing, maintaining, and interpreting the modification status of specific regions of chromatin. This review focuses on recent developments in characterizing the functional and structural links between the modification status of two DNA bases 5-methylcytosine and thymine (5-methyluracil).
Keywords: DNA 5mC oxidation; DNA demethylation; DNA enzyme; DNA methylation; DNA methyltransferase; DNA-binding protein; DNA-protein interaction; Tet dioxygenases; base excision DNA glycosylases; modification-dependent and sequence-specific transcription factors; transcription factor.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Pfaffeneder T., Hackner B., Truss M., Münzel M., Müller M., Deiml C. A., Hagemeier C., Carell T. (2011) The discovery of 5-formylcytosine in embryonic stem cell DNA. Angew. Chem. Int. Ed. Engl. 50, 7008–7012 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
