

Myosin light chains: Teaching old dogs new tricks
- PMID: 26155737
- PMCID: PMC4914028
- DOI: 10.1080/19490992.2015.1054092
Myosin light chains: Teaching old dogs new tricks
Abstract
The myosin holoenzyme is a multimeric protein complex consisting of heavy chains and light chains. Myosin light chains are calmodulin family members which are crucially involved in the mechanoenzymatic function of the myosin holoenzyme. This review examines the diversity of light chains within the myosin superfamily, discusses interactions between the light chain and the myosin heavy chain as well as regulatory and structural functions of the light chain as a subunit of the myosin holoenzyme. It covers aspects of the myosin light chain in the localization of the myosin holoenzyme, protein-protein interactions and light chain binding to non-myosin binding partners. Finally, this review challenges the dogma that myosin regulatory and essential light chain exclusively associate with conventional myosin heavy chains while unconventional myosin heavy chains usually associate with calmodulin.
Keywords: actin; calmodulin; cytoskeleton; essential light chain; live cell imaging; myosin; regulatory light chain.
Figures
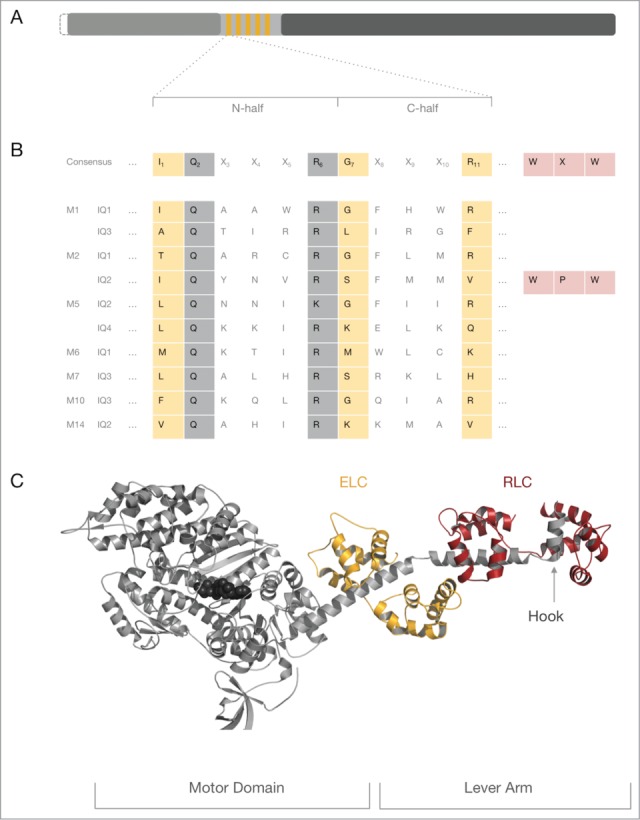
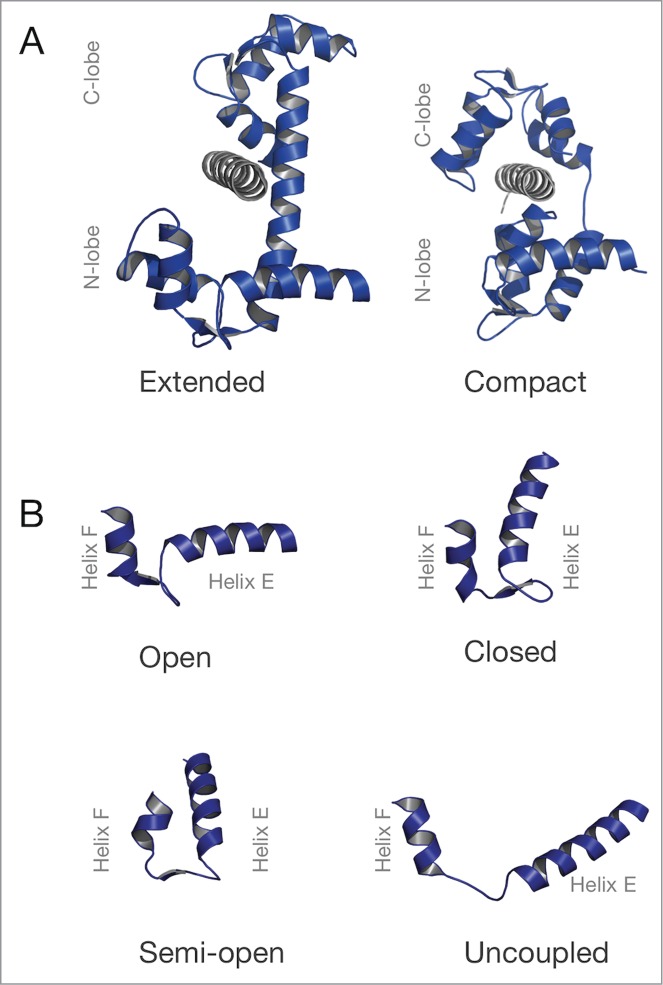
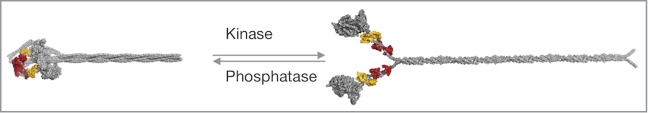
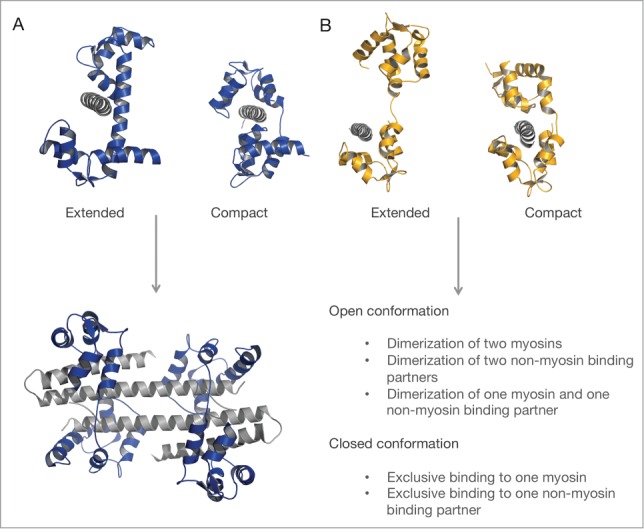
References
-
- Sellers JR. Myosins: a diverse superfamily. Biochim Biophys Acta 2000; 1496:3-22; PMID:10722873; http://dx.doi.org/10.1016/S0167-4889(00)00005-7 - DOI - PubMed
-
- Odronitz F, Kollmar M. Drawing the tree of eukaryotic life based on the analysis of 2,269 manually annotated myosins from 328 species. Genome Biol 2007; 8:R196; PMID:17877792; http://dx.doi.org/10.1186/gb-2007-8-9-r196 - DOI - PMC - PubMed
-
- Cheney RE, Mooseker MS. Unconventional myosins. Curr Opin Cell Biol 1992; 4:27-35; PMID:1558751; http://dx.doi.org/10.1016/0955-0674(92)90055-H - DOI - PubMed
-
- Bahler M, Rhoads A. Calmodulin signaling via the IQ motif. FEBS Lett 2002; 513:107-13; PMID:11911888; http://dx.doi.org/10.1016/S0014-5793(01)03239-2 - DOI - PubMed
-
- Vetter SW, Leclerc E. Novel aspects of calmodulin target recognition and activation. Eur J Biochem 2003; 270:404-14; PMID:12542690; http://dx.doi.org/10.1046/j.1432-1033.2003.03414.x - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources