

Seed shattering: from models to crops
- PMID: 26157453
- PMCID: PMC4478375
- DOI: 10.3389/fpls.2015.00476
Seed shattering: from models to crops
Abstract
Seed shattering (or pod dehiscence, or fruit shedding) is essential for the propagation of their offspring in wild plants but is a major cause of yield loss in crops. In the dicot model species, Arabidopsis thaliana, pod dehiscence necessitates a development of the abscission zones along the pod valve margins. In monocots, such as cereals, an abscission layer in the pedicle is required for the seed shattering process. In the past decade, great advances have been made in characterizing the genetic contributors that are involved in the complex regulatory network in the establishment of abscission cell identity. We summarize the recent burgeoning progress in the field of genetic regulation of pod dehiscence and fruit shedding, focusing mainly on the model species A. thaliana with its close relatives and the fleshy fruit species tomato, as well as the genetic basis responsible for the parallel loss of seed shattering in domesticated crops. This review shows how these individual genes are co-opted in the developmental process of the tissues that guarantee seed shattering. Research into the genetic mechanism underlying seed shattering provides a premier prerequisite for the future breeding program for harvest in crops.
Keywords: domestication; domestication syndrome; fruit shedding; genetic regulation; indehiscent fruit; pod dehiscence; seed shattering.
Figures
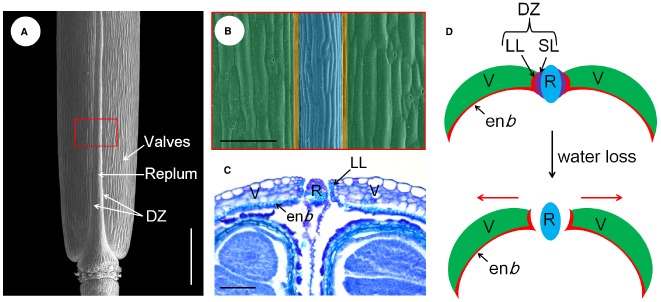
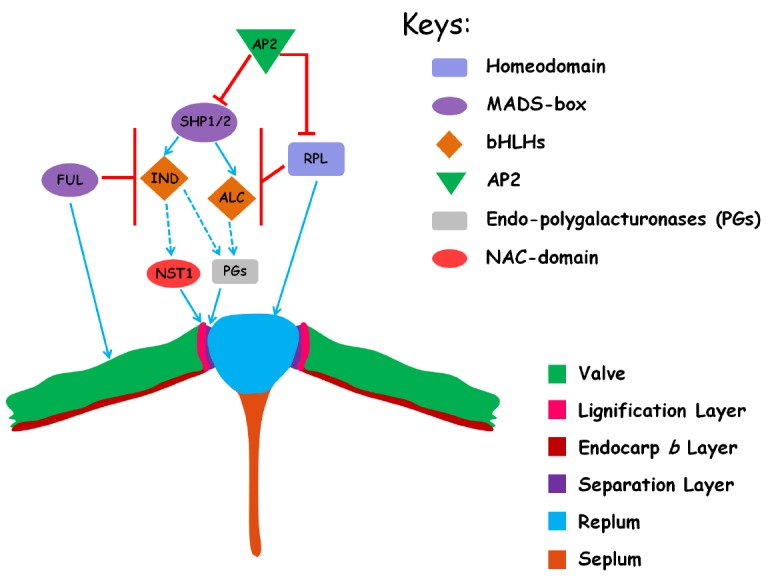
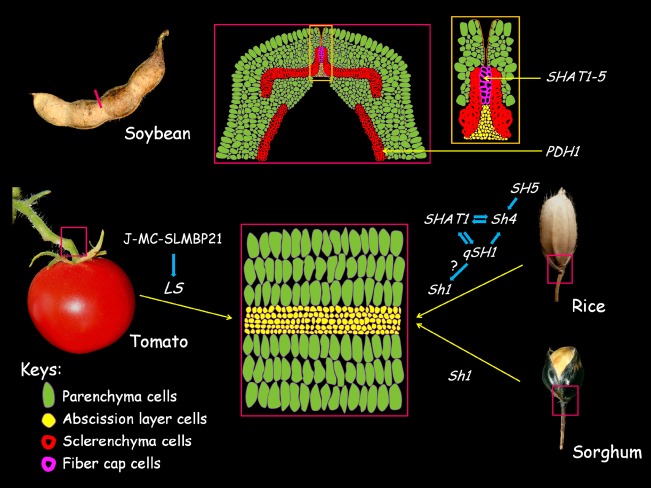
References
-
- Appel O., Al-Shehbaz I. A. (2003). Cruciferae. Flowering Plants–Dicotyledons. pp. 75–174. Berlin: Springer.
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous