

All-optical mapping of barrel cortex circuits based on simultaneous voltage-sensitive dye imaging and channelrhodopsin-mediated photostimulation
- PMID: 26158003
- PMCID: PMC4478985
- DOI: 10.1117/1.NPh.2.2.021013
All-optical mapping of barrel cortex circuits based on simultaneous voltage-sensitive dye imaging and channelrhodopsin-mediated photostimulation
Abstract
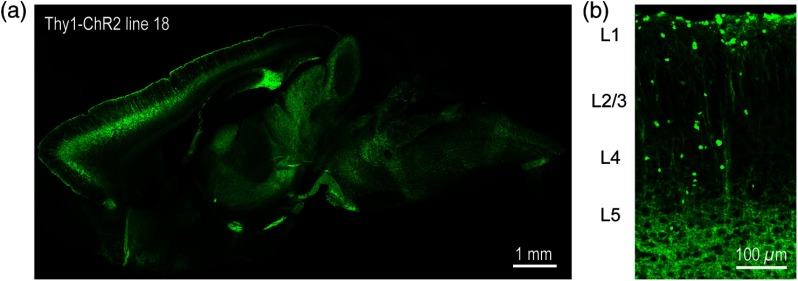
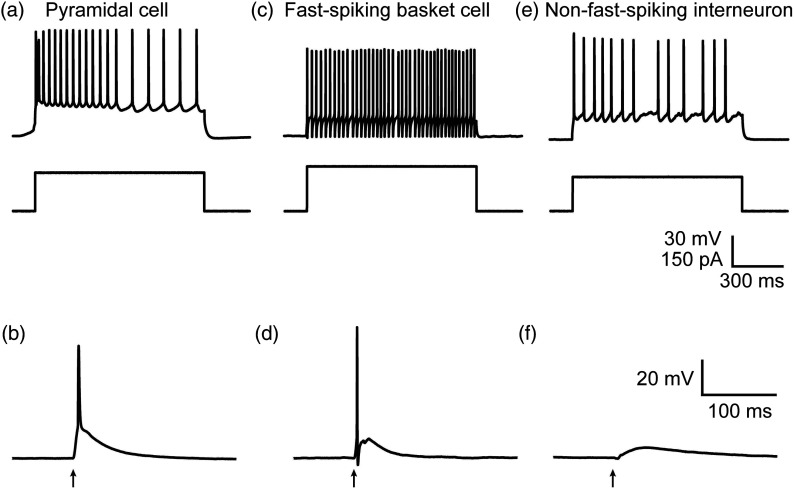
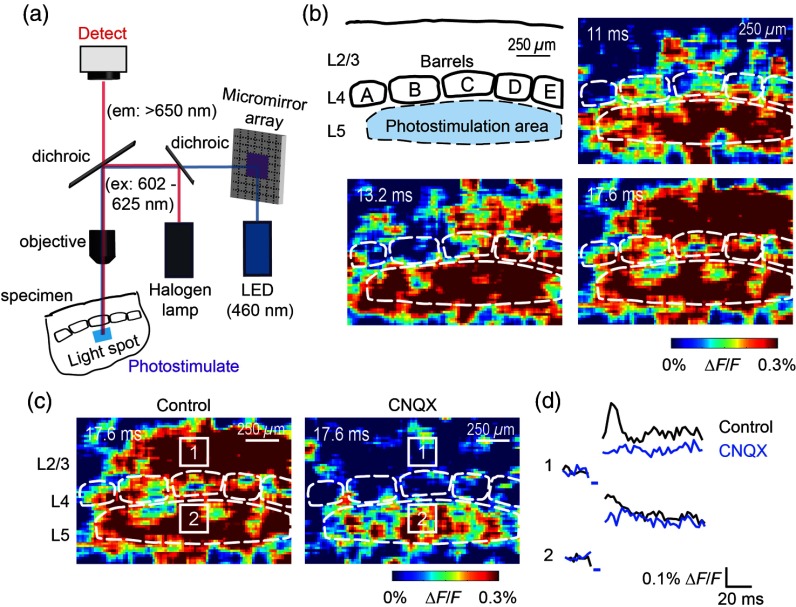
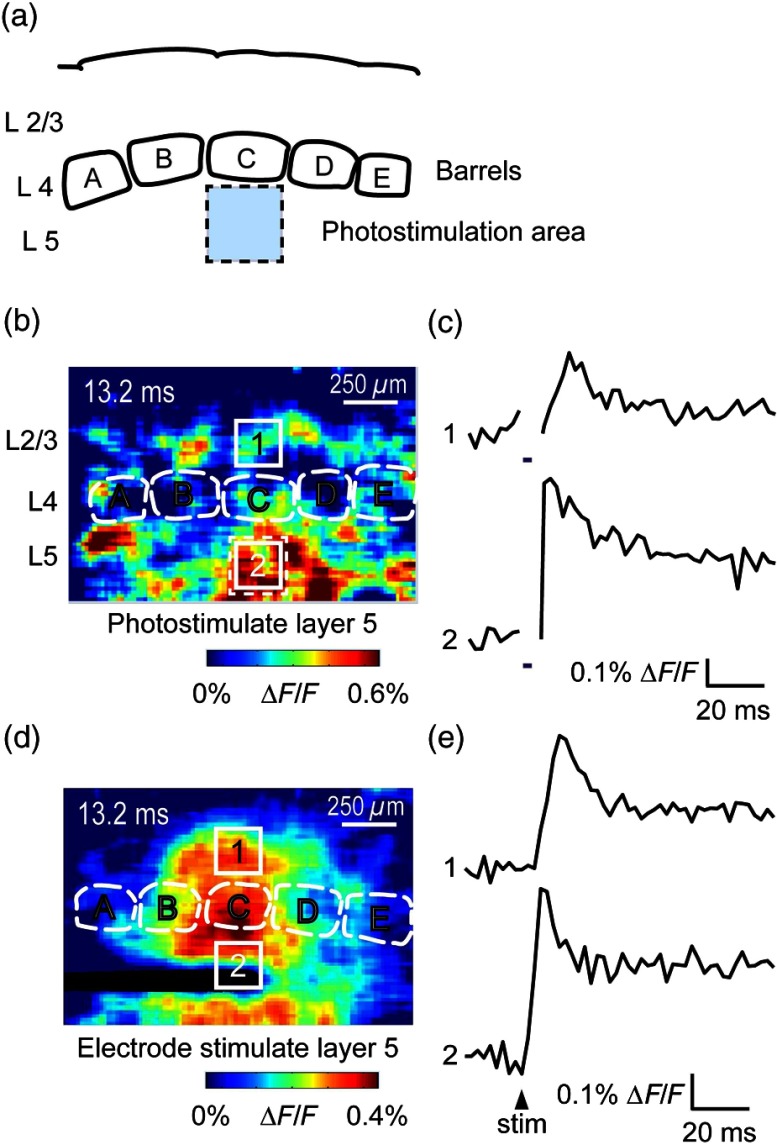
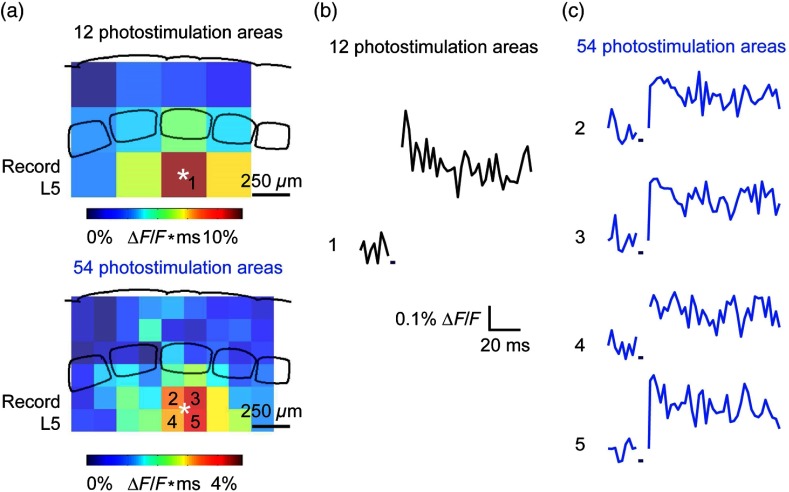
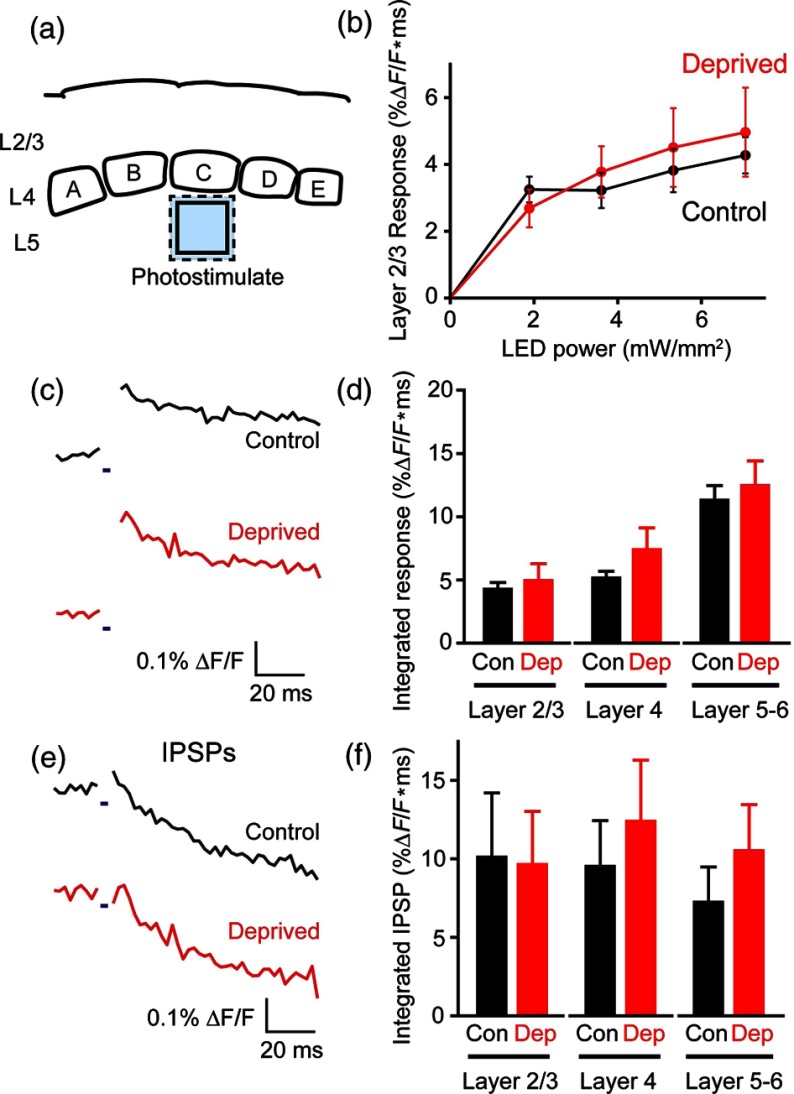
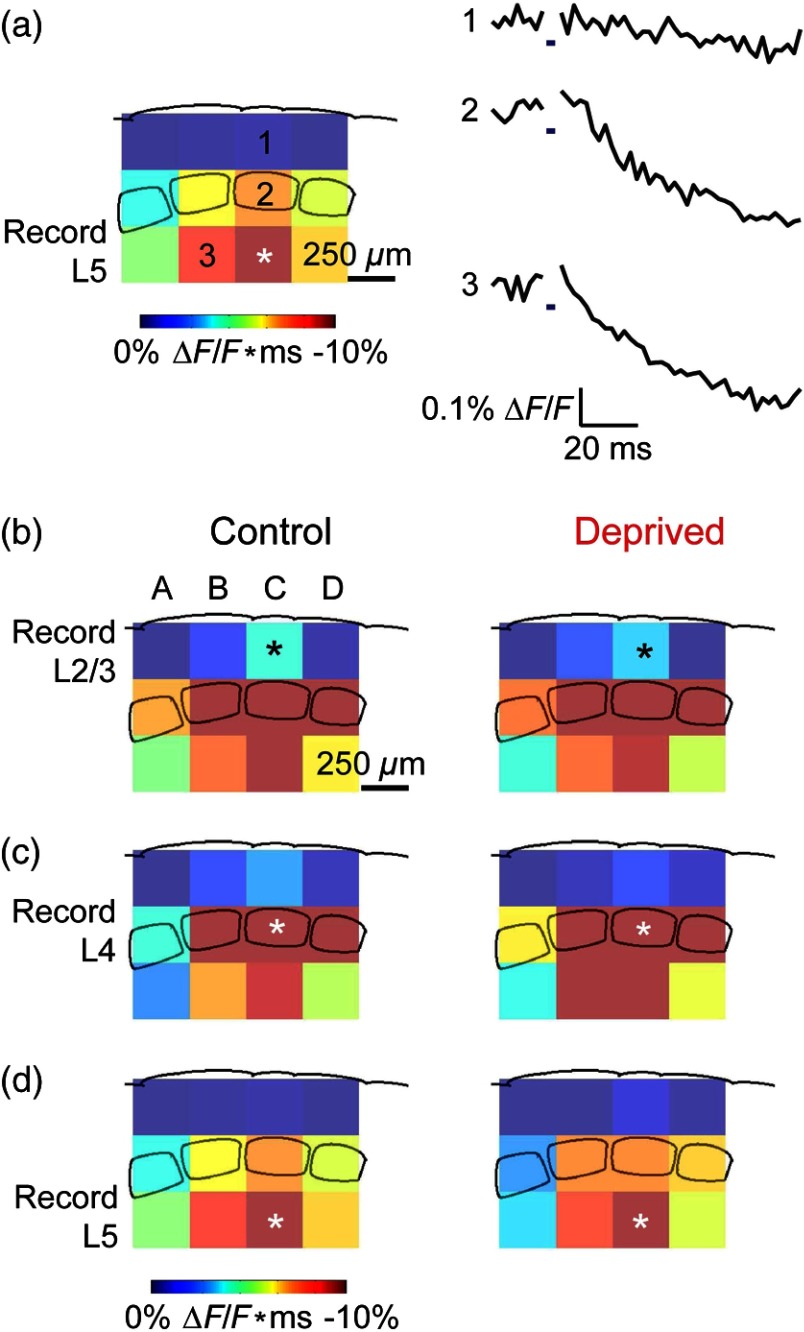
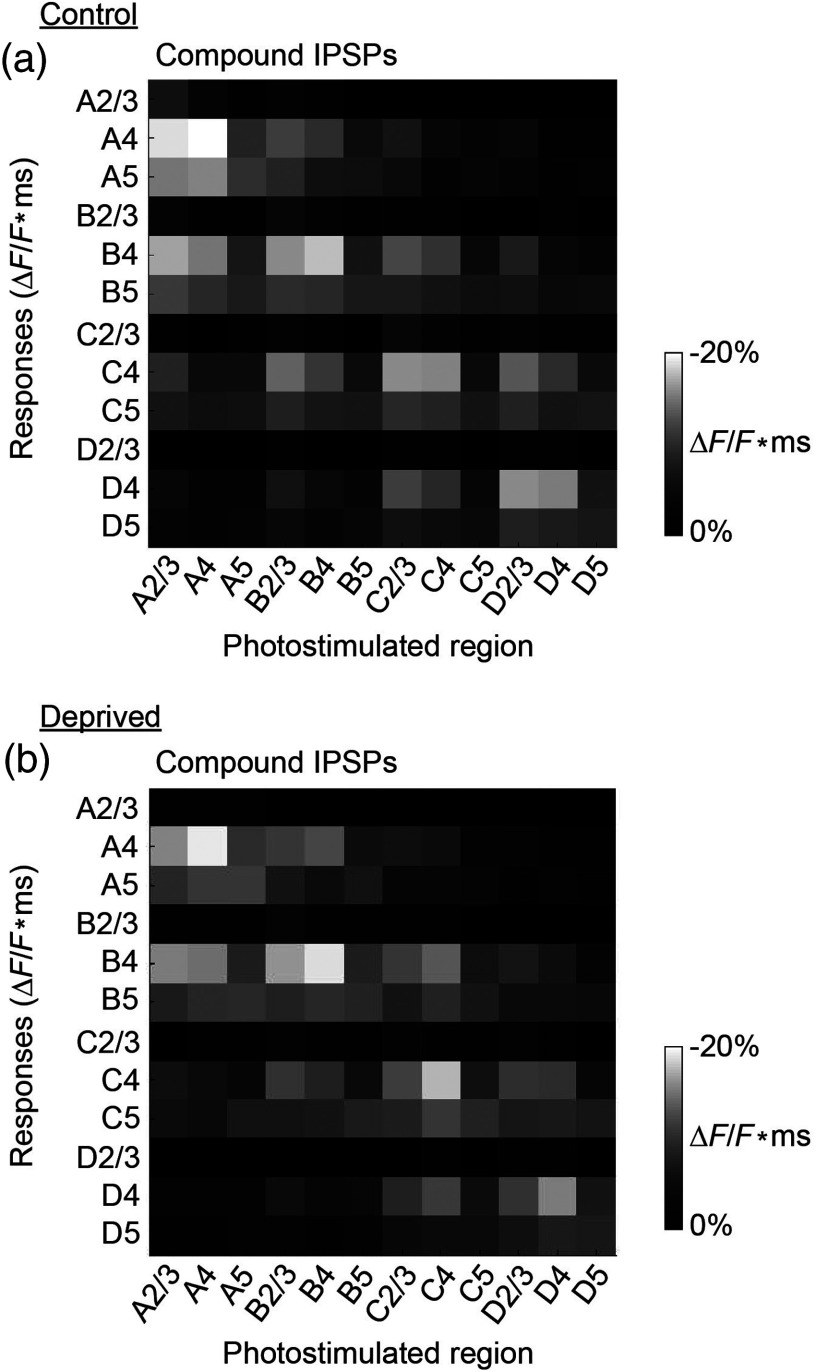
We describe an experimental approach that uses light to both control and detect neuronal activity in mouse barrel cortex slices: blue light patterned by a digital micromirror array system allowed us to photostimulate specific layers and columns, while a red-shifted voltage-sensitive dye was used to map out large-scale circuit activity. We demonstrate that such all-optical mapping can interrogate various circuits in somatosensory cortex by sequentially activating different layers and columns. Further, mapping in slices from whisker-deprived mice demonstrated that chronic sensory deprivation did not significantly alter feedforward inhibition driven by layer 5 pyramidal neurons. Further development of voltage-sensitive optical probes should allow this all-optical mapping approach to become an important and high-throughput tool for mapping circuit interactions in the brain.
Keywords: barrel cortex; channelrhodopsin; optogenetics; voltage-sensitive dye.
Figures
Similar articles
-
In vivo voltage-sensitive dye imaging of mouse cortical activity with mesoscopic optical tomography.Neurophotonics. 2020 Oct;7(4):041402. doi: 10.1117/1.NPh.7.4.041402. Epub 2020 Dec 2. Neurophotonics. 2020. PMID: 33274250 Free PMC article.
-
Probing the function of neuronal populations: combining micromirror-based optogenetic photostimulation with voltage-sensitive dye imaging.Neurosci Res. 2013 Jan;75(1):76-81. doi: 10.1016/j.neures.2012.11.006. Epub 2012 Dec 17. Neurosci Res. 2013. PMID: 23254260 Free PMC article.
-
Study of the cortical representation of whisker frequency selectivity using voltage-sensitive dye optical imaging.Intravital. 2016 Feb 18;5(1):e1142637. doi: 10.1080/21659087.2016.1142637. eCollection 2016. Intravital. 2016. PMID: 28243518 Free PMC article.
-
Mapping functional connectivity in barrel-related columns reveals layer- and cell type-specific microcircuits.Brain Struct Funct. 2007 Sep;212(2):107-19. doi: 10.1007/s00429-007-0147-z. Epub 2007 Jun 26. Brain Struct Funct. 2007. PMID: 17717691 Review.
-
All-optical interrogation of neural circuits in behaving mice.Nat Protoc. 2022 Jul;17(7):1579-1620. doi: 10.1038/s41596-022-00691-w. Epub 2022 Apr 27. Nat Protoc. 2022. PMID: 35478249 Free PMC article. Review.
Cited by
-
Motor cortex stimulation ameliorates parkinsonian locomotor deficits: effectual and mechanistic differences from subthalamic modulation.NPJ Parkinsons Dis. 2025 Feb 19;11(1):32. doi: 10.1038/s41531-025-00879-3. NPJ Parkinsons Dis. 2025. PMID: 39971974 Free PMC article.
-
Defining a critical period for inhibitory circuits within the somatosensory cortex.Sci Rep. 2017 Aug 4;7(1):7271. doi: 10.1038/s41598-017-07400-8. Sci Rep. 2017. PMID: 28779074 Free PMC article.
-
Mammalian cortical voltage imaging using genetically encoded voltage indicators: a review honoring professor Amiram Grinvald.Neurophotonics. 2017 Jul;4(3):031214. doi: 10.1117/1.NPh.4.3.031214. Epub 2017 May 4. Neurophotonics. 2017. PMID: 28491906 Free PMC article. Review.
-
In vivo voltage-sensitive dye imaging of mouse cortical activity with mesoscopic optical tomography.Neurophotonics. 2020 Oct;7(4):041402. doi: 10.1117/1.NPh.7.4.041402. Epub 2020 Dec 2. Neurophotonics. 2020. PMID: 33274250 Free PMC article.
-
Optogenetic cortical spreading depression originating from the primary visual cortex induces migraine-like pain and anxiety behaviors in freely moving C57BL/6 J mice.J Headache Pain. 2025 Feb 26;26(1):44. doi: 10.1186/s10194-025-01983-8. J Headache Pain. 2025. PMID: 40011818 Free PMC article.
