

Activation of Human Toll-like Receptor 4 (TLR4)·Myeloid Differentiation Factor 2 (MD-2) by Hypoacylated Lipopolysaccharide from a Clinical Isolate of Burkholderia cenocepacia
- PMID: 26160169
- PMCID: PMC4571861
- DOI: 10.1074/jbc.M115.649087
Activation of Human Toll-like Receptor 4 (TLR4)·Myeloid Differentiation Factor 2 (MD-2) by Hypoacylated Lipopolysaccharide from a Clinical Isolate of Burkholderia cenocepacia
Abstract
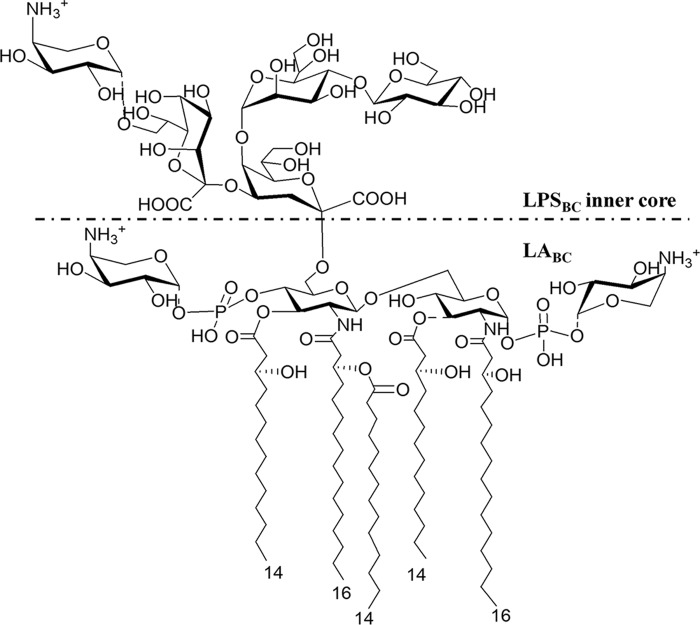
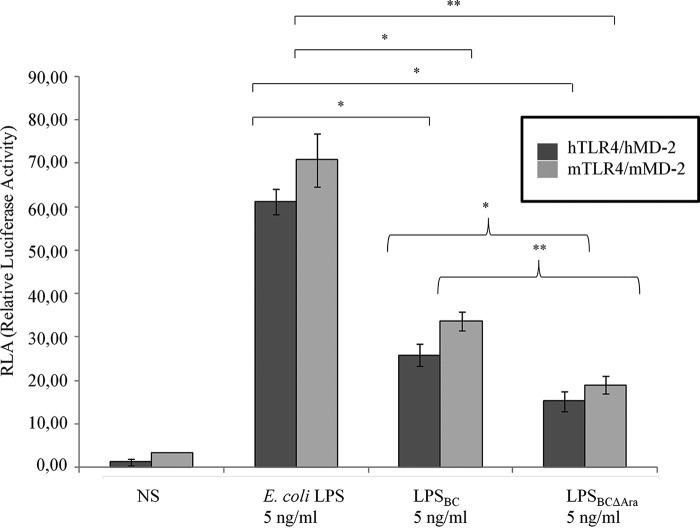
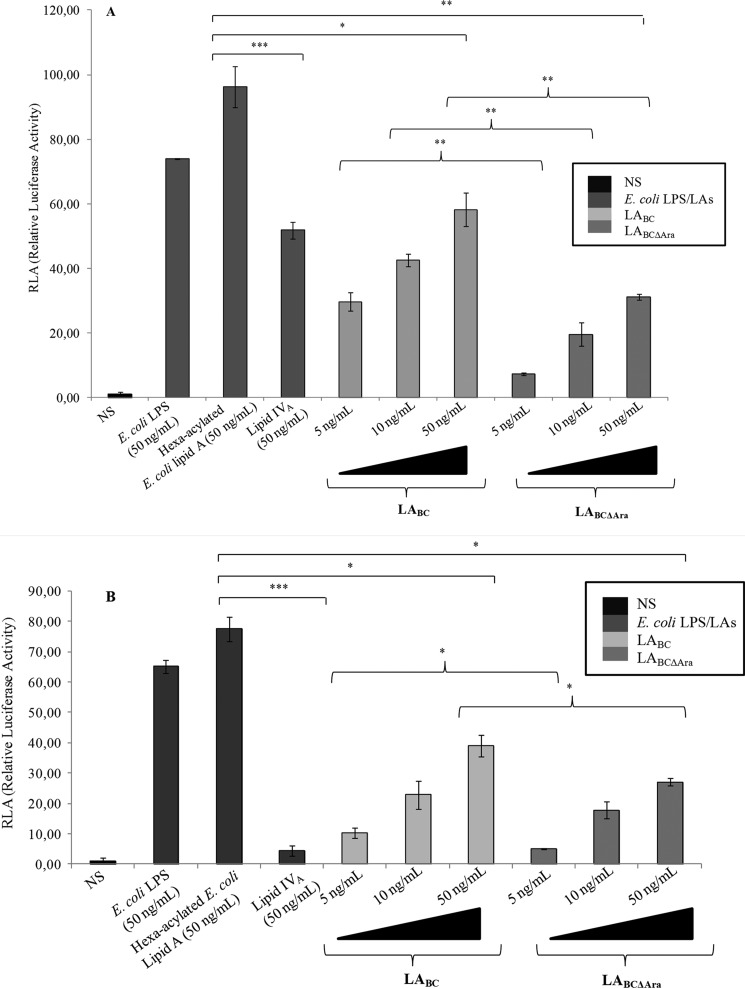
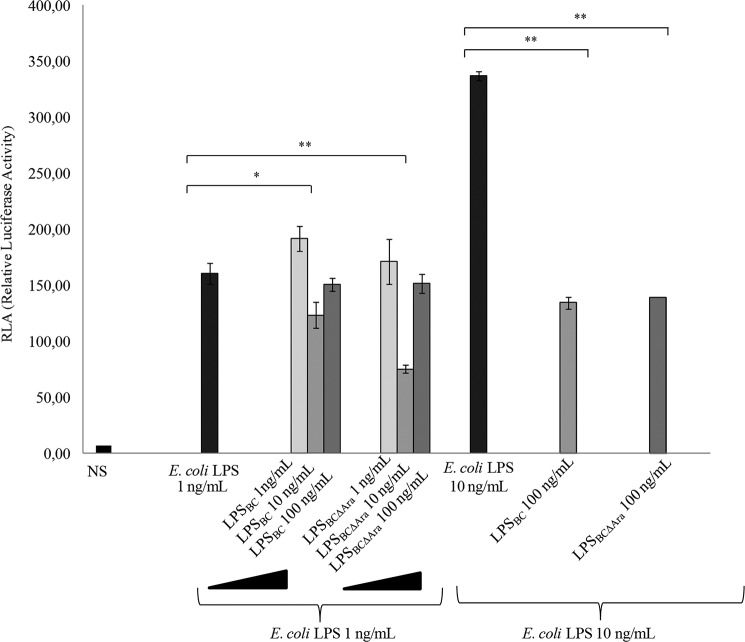
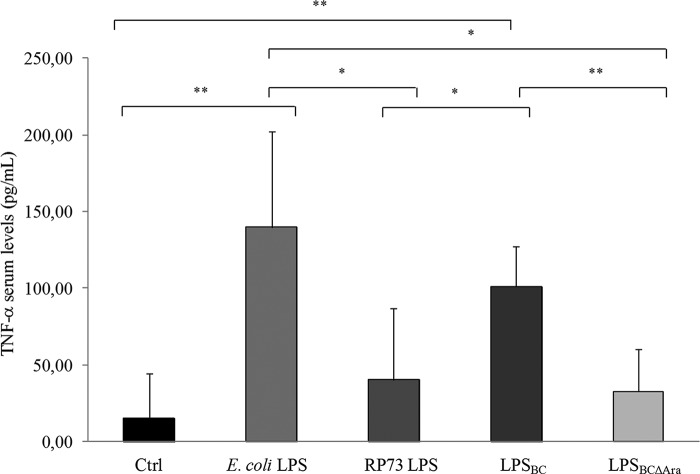
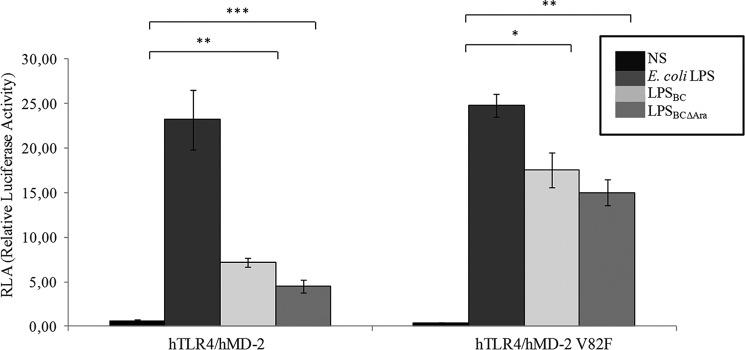
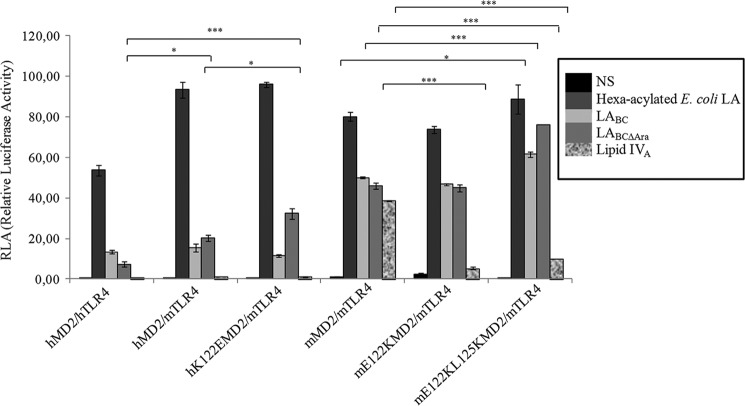
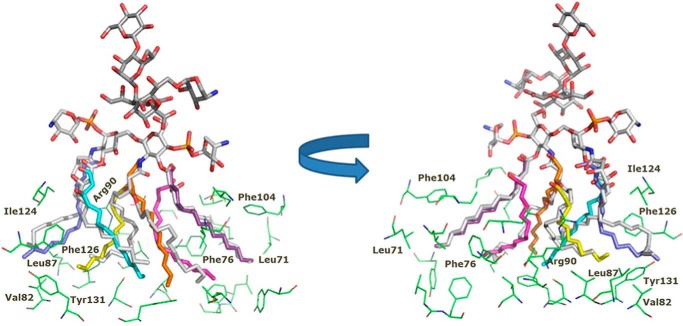
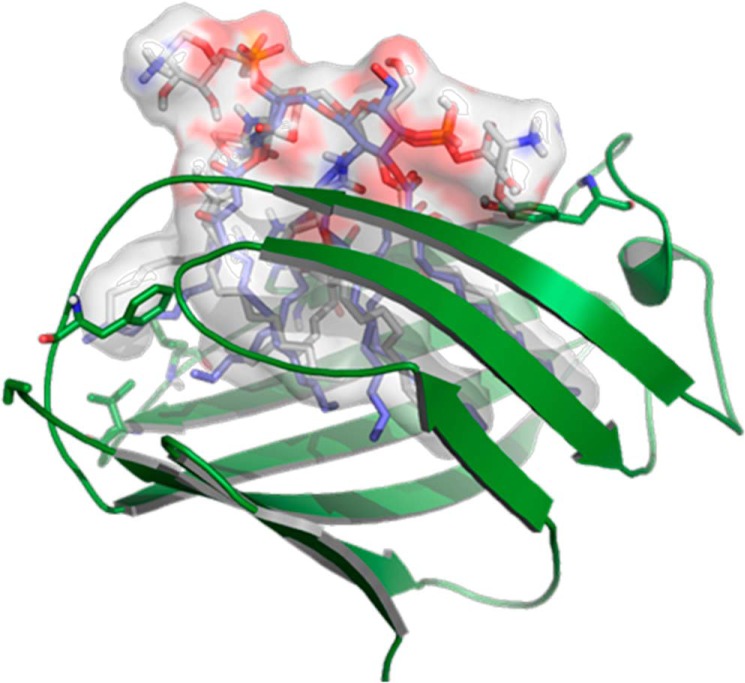
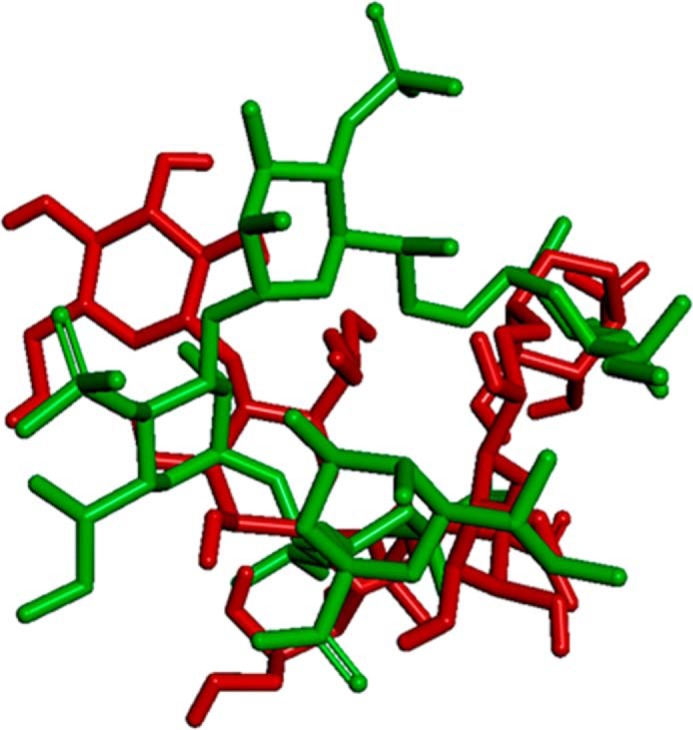
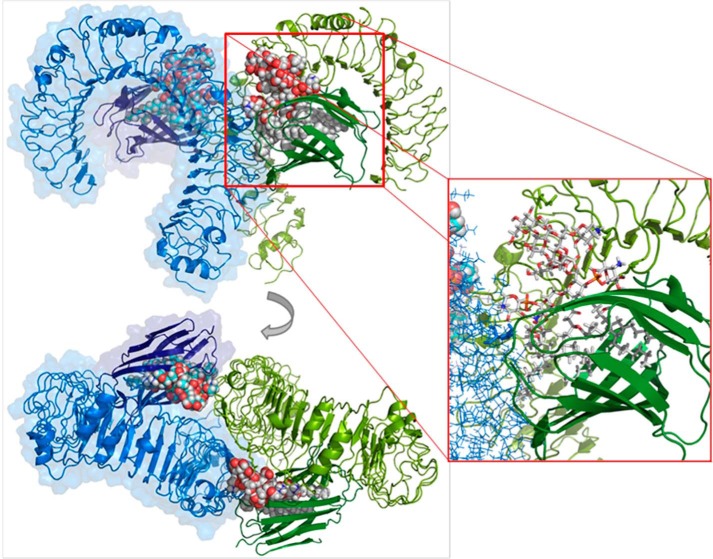
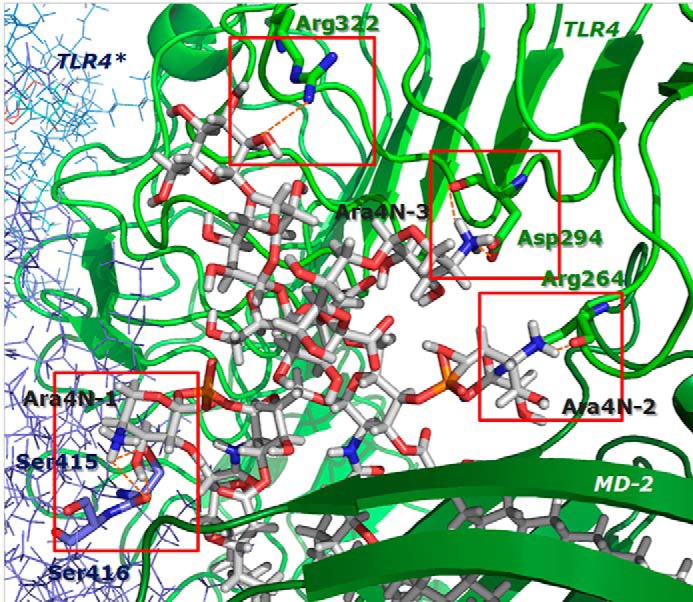
Lung infection by Burkholderia species, in particular Burkholderia cenocepacia, accelerates tissue damage and increases post-lung transplant mortality in cystic fibrosis patients. Host-microbe interplay largely depends on interactions between pathogen-specific molecules and innate immune receptors such as Toll-like receptor 4 (TLR4), which recognizes the lipid A moiety of the bacterial lipopolysaccharide (LPS). The human TLR4·myeloid differentiation factor 2 (MD-2) LPS receptor complex is strongly activated by hexa-acylated lipid A and poorly activated by underacylated lipid A. Here, we report that B. cenocepacia LPS strongly activates human TLR4·MD-2 despite its lipid A having only five acyl chains. Furthermore, we show that aminoarabinose residues in lipid A contribute to TLR4-lipid A interactions, and experiments in a mouse model of LPS-induced endotoxic shock confirmed the proinflammatory potential of B. cenocepacia penta-acylated lipid A. Molecular modeling combined with mutagenesis of TLR4-MD-2 interactive surfaces suggests that longer acyl chains and the aminoarabinose residues in the B. cenocepacia lipid A allow exposure of the fifth acyl chain on the surface of MD-2 enabling interactions with TLR4 and its dimerization. Our results provide a molecular model for activation of the human TLR4·MD-2 complex by penta-acylated lipid A explaining the ability of hypoacylated B. cenocepacia LPS to promote proinflammatory responses associated with the severe pathogenicity of this opportunistic bacterium.
Keywords: Burkholderia; Gram-negative bacteria; TLR4/MD-2; cystic fibrosis; innate immunity; lipopolysaccharide (LPS); molecular modeling.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

References
-
- Janeway C. A., Jr., Medzhitov R. (2002) Innate immune recognition. Annu. Rev. Immunol. 20, 197–216 - PubMed
-
- Gay N. J., Gangloff M. (2007) Structure and function of Toll receptors and their ligands. Annu. Rev. Biochem. 76, 141–165 - PubMed
-
- Broz P., Monack D. M. (2013) Newly described pattern recognition receptors team up against intracellular pathogens. Nat. Rev. Immunol. 13, 551–565 - PubMed
-
- Nagai Y., Akashi S., Nagafuku M., Ogata M., Iwakura Y., Akira S., Kitamura T., Kosugi A., Kimoto M., Miyake K. (2002) Essential role of MD-2 in LPS responsiveness and TLR4 distribution. Nat. Immunol. 3, 667–672 - PubMed
-
- Angus D. C., Linde-Zwirble W. T., Lidicker J., Clermont G., Carcillo J., Pinsky M. R. (2001) Epidemiology of severe sepsis in the United States: analysis of incidence, outcome, and associated costs of care. Crit. Care Med. 29, 1303–1310 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
