

A 45-Amino-Acid Scaffold Mined from the PDB for High-Affinity Ligand Engineering
- PMID: 26165154
- PMCID: PMC4536934
- DOI: 10.1016/j.chembiol.2015.06.012
A 45-Amino-Acid Scaffold Mined from the PDB for High-Affinity Ligand Engineering
Abstract
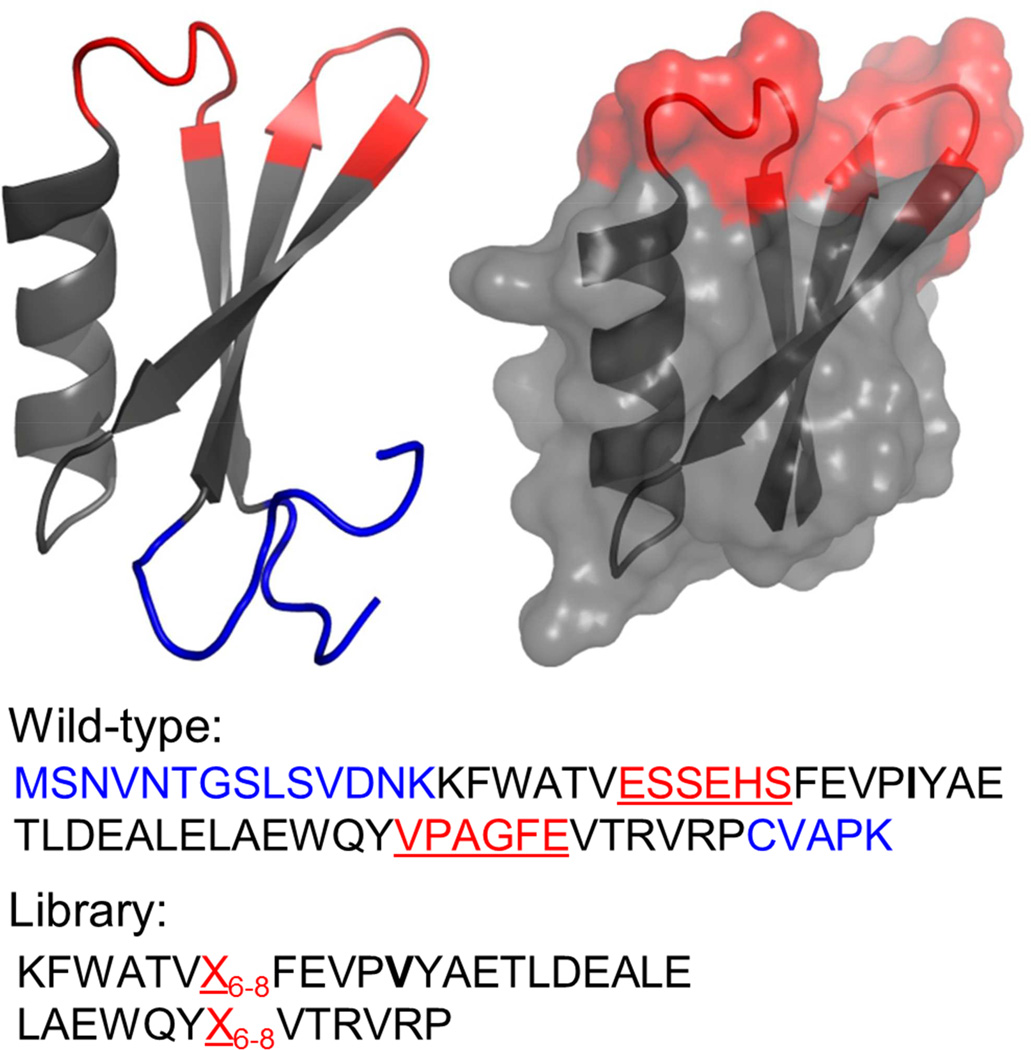
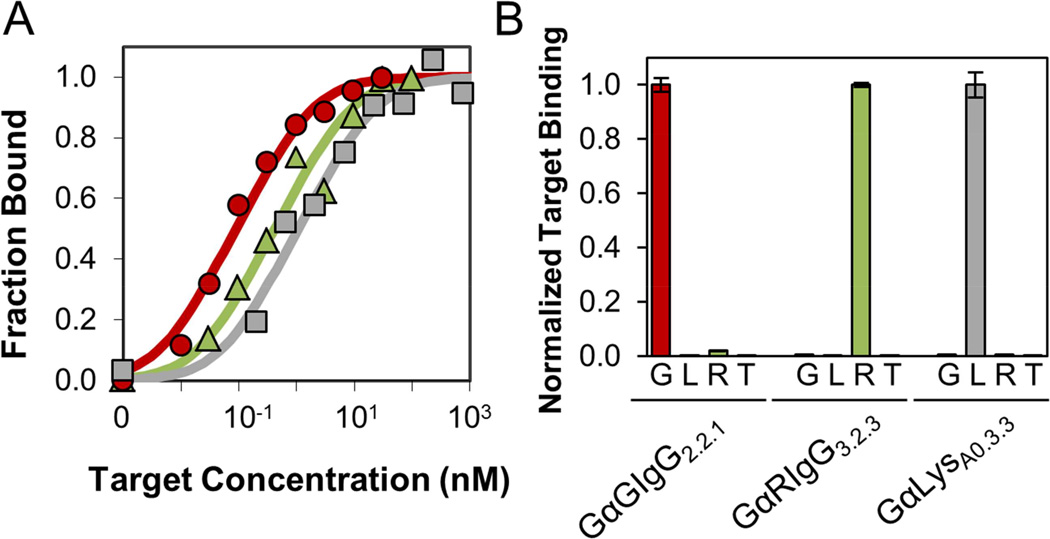
Small protein ligands can provide superior physiological distribution compared with antibodies, and improved stability, production, and specific conjugation. Systematic evaluation of the PDB identified a scaffold to push the limits of small size and robust evolution of stable, high-affinity ligands: 45-residue T7 phage gene 2 protein (Gp2) contains an α helix opposite a β sheet with two adjacent loops amenable to mutation. De novo ligand discovery from 10(8) mutants and directed evolution toward four targets yielded target-specific binders with affinities as strong as 200 ± 100 pM, Tms from 65 °C ± 3 °C to 80°C ± 1 °C, and retained activity after thermal denaturation. For cancer targeting, a Gp2 domain for epidermal growth factor receptor was evolved with 18 ± 8 nM affinity, receptor-specific binding, and high thermal stability with refolding. The efficiency of evolving new binding function and the size, affinity, specificity, and stability of evolved domains render Gp2 a uniquely effective ligand scaffold.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures




References
-
- Ackerman SE, Currier NV, Bergen JM, Cochran JR. Cystine-knot peptides: emerging tools for cancer imaging and therapy. Expert Rev. Proteomics. 2014;11:561–572. - PubMed
-
- Banta S, Dooley K, Shur O. Replacing antibodies: engineering new binding proteins. Annu. Rev. Biomed. Eng. 2013;15:93–113. - PubMed
-
- Baum RP, Prasad V, Müller D, Schuchardt C, Orlova A, Wennborg A, Tolmachev V, Feldwisch J. Molecular imaging of HER2-expressing malignant tumors in breast cancer patients using synthetic 111In- or 68Ga-labeled affibody molecules. J. Nucl. Med. 2010;51:892–897. - PubMed
-
- Binz HK, Amstutz P, Plückthun A. Engineering novel binding proteins from nonimmunoglobulin domains. Nat. Biotechnol. 2005;23:1257–1268. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
