

Integrin α4β7 Expression Increases HIV Susceptibility in Activated Cervical CD4+ T Cells by an HIV Attachment-Independent Mechanism
- PMID: 26167616
- PMCID: PMC4503378
- DOI: 10.1097/QAI.0000000000000676
Integrin α4β7 Expression Increases HIV Susceptibility in Activated Cervical CD4+ T Cells by an HIV Attachment-Independent Mechanism
Abstract
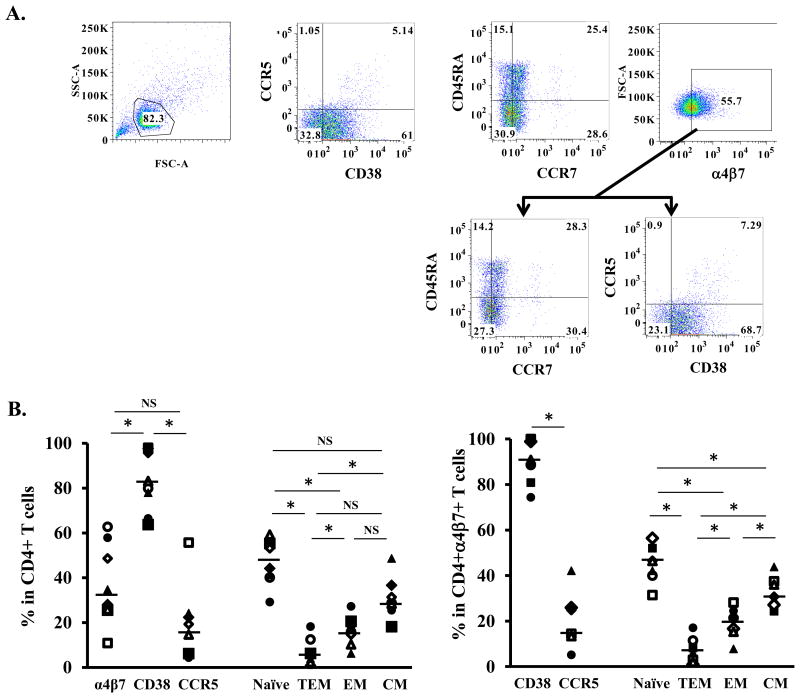
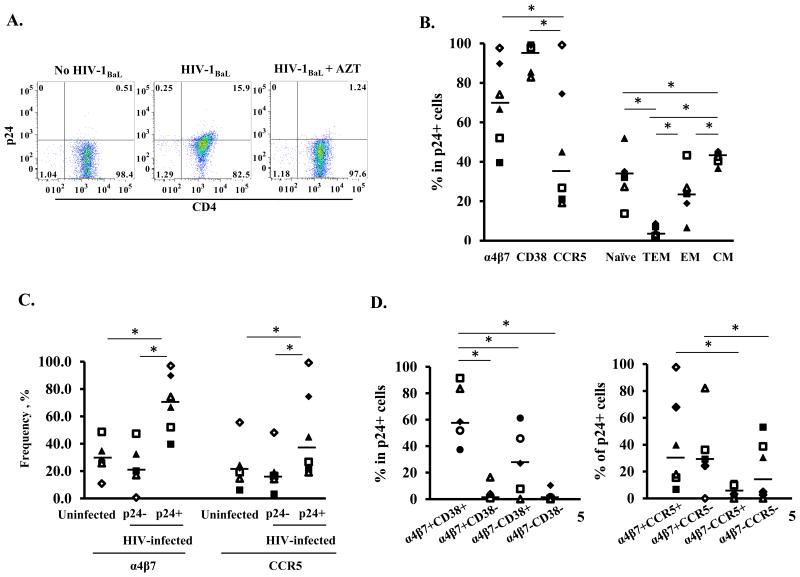
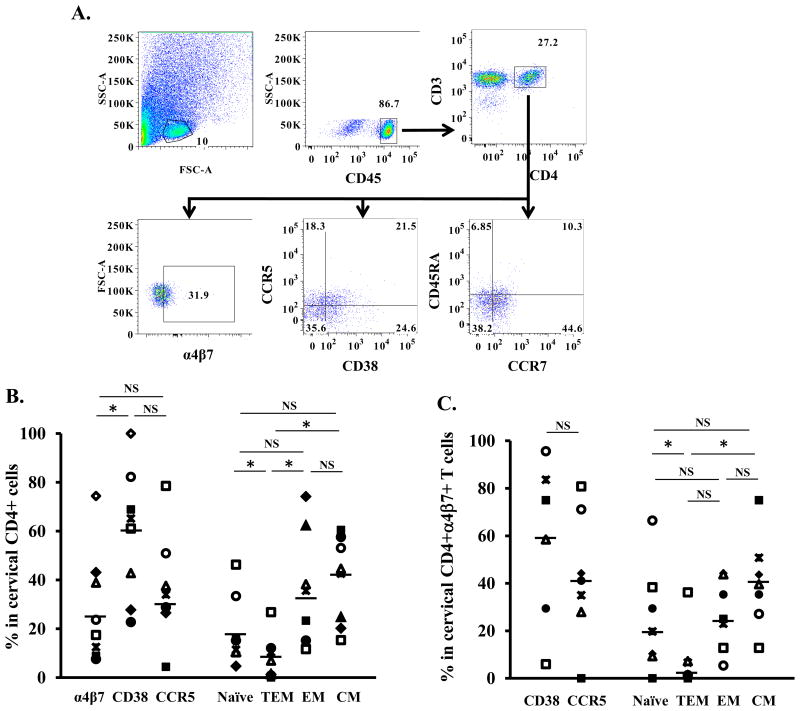
Background: CD4 T cells are crucial for the establishment and dissemination of HIV in mucosal tissues during acute infection. Studies indicate that integrin α4β7 CD4 T cells are preferentially infected by HIV in vitro and during acute SIV infection. The integrin α4β7 is thought to promote HIV capture by target cells; however, the role of integrin α4β7 in HIV transmission remains controversial. In this study, we characterized immune phenotypes of human cervical T cells and examined HIV preference in integrin α4β7 CD4 T cells. In vitro all-trans retinoic acid-differentiated peripheral CD4 T cells (atRA-differentiated cells) were included as a comparison.
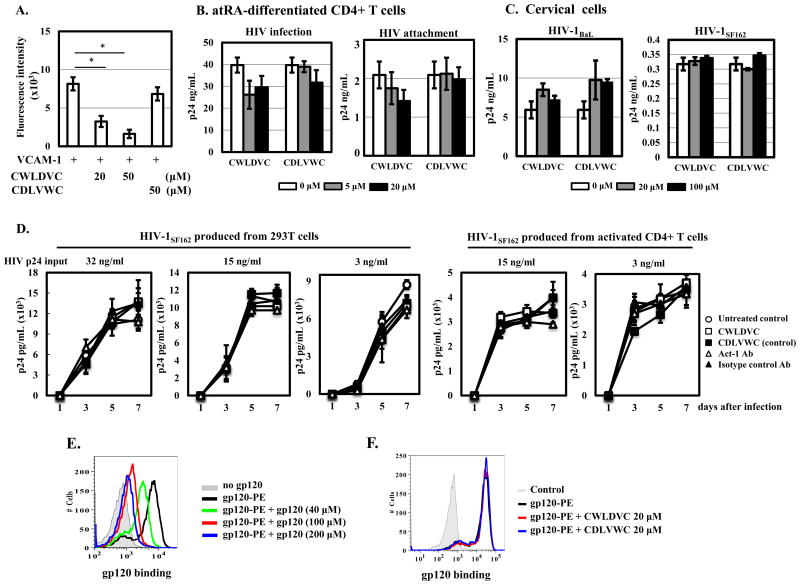
Results: In both peripheral and cervical cells, the majority of HIV p24 cells were activated CD4 T cells expressing integrin α4β7. Among infected atRA-differentiated cells, the frequency of CCR5 expression was higher in HIV p24 cells than in HIV p24 cells; no such difference was observed in cervical cells. Neither the cyclic hexapeptide CWLDVC nor a monoclonal antibody against integrin α4β7 blocked HIV attachment or gp120 binding to target cells regardless of the presence of CD4, indicating that integrin α4β7 did not facilitate HIV capture by target cells.
Conclusions: Integrin α4β7 expression increases HIV susceptibility, but the mechanism is not through promoting HIV binding to target cells.
Figures

References
-
- Haase AT. Early events in sexual transmission of HIV and SIV and opportunities for interventions. Annu Rev Med. 2011 Feb 18;62:127–39. - PubMed
-
- Arthos J, Cicala C, Martinelli E, et al. HIV-1 envelope protein binds to and signals through integrin alpha4beta7, the gut mucosal homing receptor for peripheral T cells. Nat Immunol. 2008 Mar;9(3):301–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
