

17β-Estradiol Ameliorates Tight Junction Disruption via Repression of MMP Transcription
- PMID: 26168035
- PMCID: PMC5414681
- DOI: 10.1210/ME.2015-1124
17β-Estradiol Ameliorates Tight Junction Disruption via Repression of MMP Transcription
Abstract
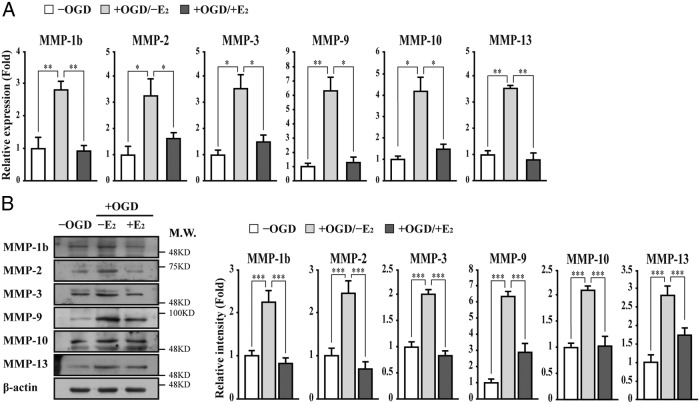
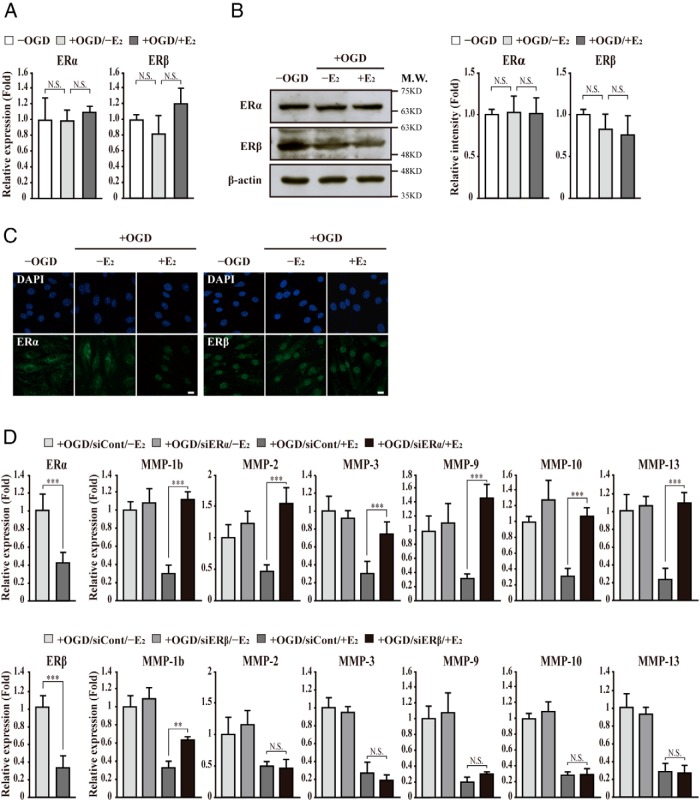
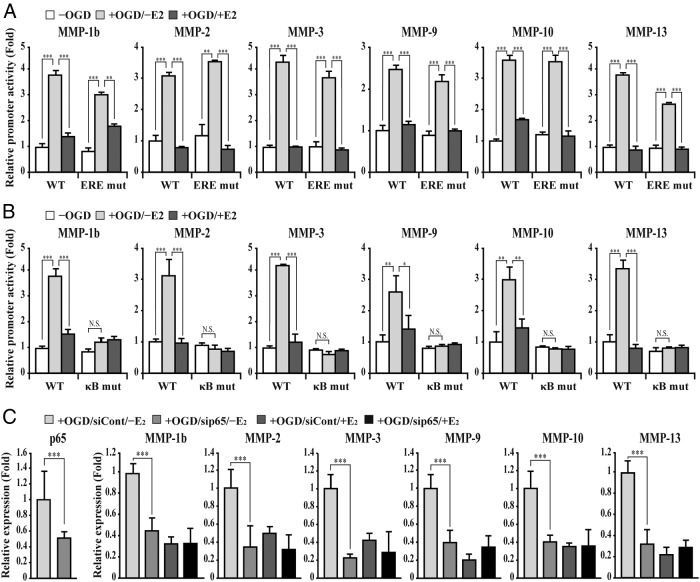
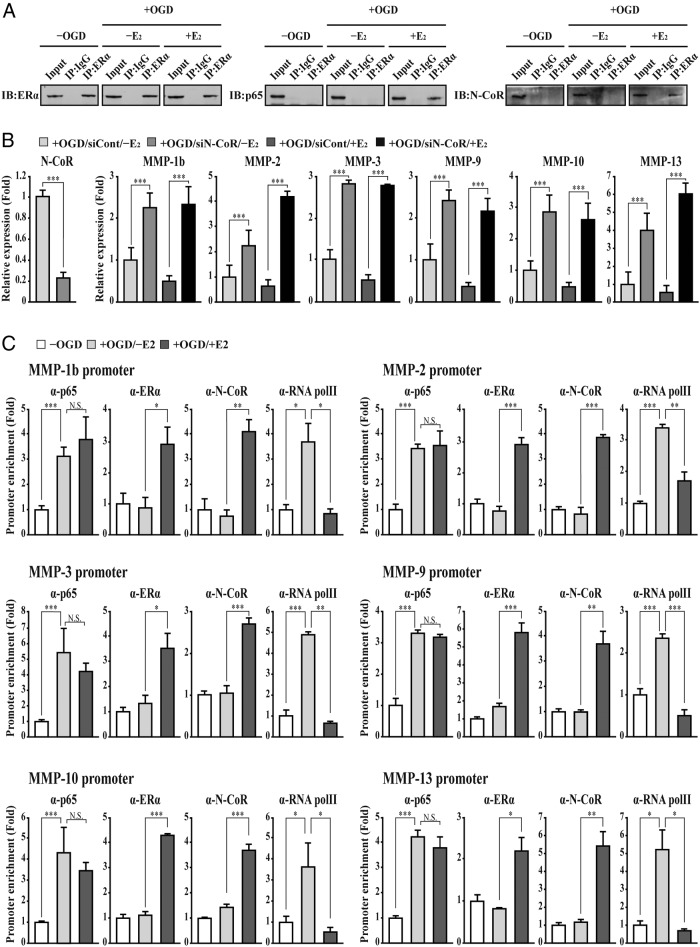
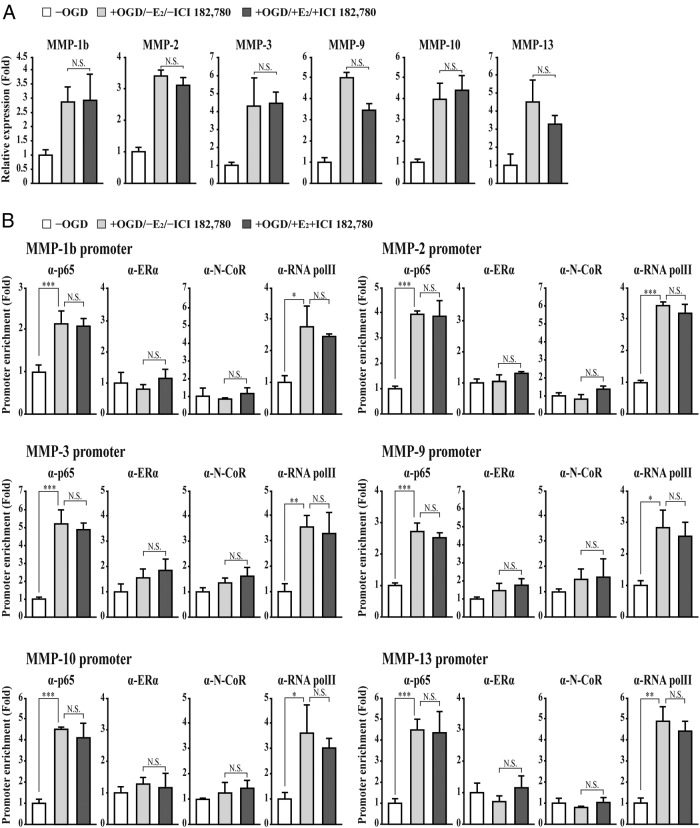
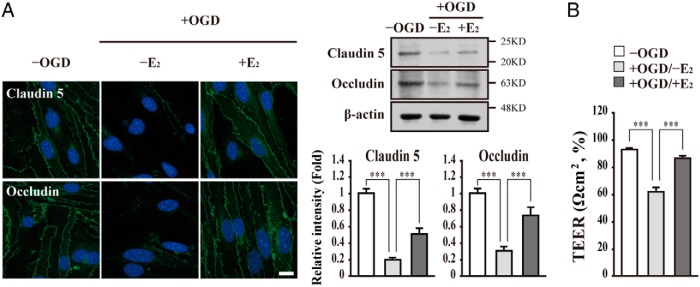
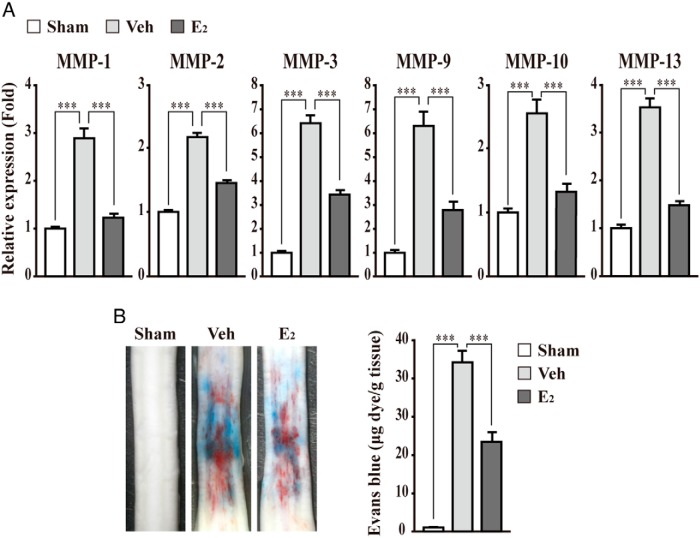
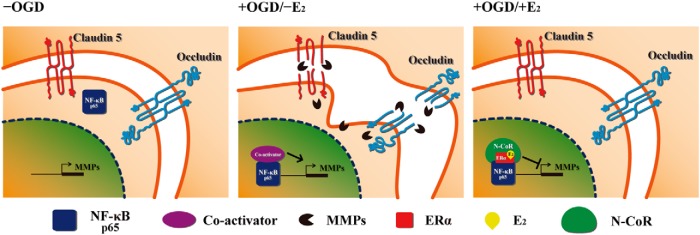
The blood-brain barrier (BBB) or blood-spinal cord barrier (BSCB) formed by capillary endothelial cells provides a physical wall between the central nervous system (CNS) and circulating blood with highly selective permeability. BBB/BSCB disruption by activation of matrix metalloproteinases (MMPs) has been shown to result in further neurological damage after CNS injury. Recently it has been discovered that estrogen attenuates BBB/BSCB disruption in in vitro and in vivo models. However, the molecular mechanism underlying the estrogen-mediated attenuation of BBB/BSCB disruption has not been elucidated fully. In the present study, we found that 17β-estradiol (E2) suppresses nuclear factor-κB-dependent MMP-1b, MMP-2, MMP-3, MMP-9, MMP-10, and MMP-13 gene activation in microvessel endothelial bEnd.3 cells subjected to oxygen and glucose deprivation/reperfusion injury. E2 induced the recruitment of ERα and nuclear receptor corepressor to the nuclear factor-κB binding site on the MMPs' gene promoters. Consistently, ER antagonist ICI 182.780 showed opposite effects of E2. We further found that E2 attenuates tight junction disruption through the decreased degradation of tight junction proteins in bEnd.3 cells subjected to oxygen and glucose deprivation-reperfusion injury. In addition, E2 suppressed the up-regulation of MMP expression, leading to a decreased BSCB disruption in the injured spinal cord. In conclusion, we discovered the molecular mechanism underlying the protective role of estrogenin BBB/BSCB disruption using an in vitro and in vivo model. Our study suggests that estrogens may provide a potential therapeutic intervention for preserving BBB/BSCB integrity after CNS injury.
Figures
Similar articles
-
17β-estradiol inhibits MMP-9 and SUR1/TrpM4 expression and activation and thereby attenuates BSCB disruption/hemorrhage after spinal cord injury in male rats.Endocrinology. 2015 May;156(5):1838-50. doi: 10.1210/en.2014-1832. Epub 2015 Mar 12. Endocrinology. 2015. PMID: 25763638
-
Matrix metalloproteinase-3 promotes early blood-spinal cord barrier disruption and hemorrhage and impairs long-term neurological recovery after spinal cord injury.Am J Pathol. 2014 Nov;184(11):2985-3000. doi: 10.1016/j.ajpath.2014.07.016. Epub 2014 Oct 14. Am J Pathol. 2014. PMID: 25325922
-
Activation of classical estrogen receptor subtypes reduces tight junction disruption of brain endothelial cells under ischemia/reperfusion injury.Free Radic Biol Med. 2016 Mar;92:78-89. doi: 10.1016/j.freeradbiomed.2016.01.010. Epub 2016 Jan 16. Free Radic Biol Med. 2016. PMID: 26784014
-
Propitious Therapeutic Modulators to Prevent Blood-Spinal Cord Barrier Disruption in Spinal Cord Injury.Mol Neurobiol. 2017 Jul;54(5):3578-3590. doi: 10.1007/s12035-016-9910-6. Epub 2016 May 18. Mol Neurobiol. 2017. PMID: 27194298 Review.
-
Interaction of free radicals, matrix metalloproteinases and caveolin-1 impacts blood-brain barrier permeability.Front Biosci (Schol Ed). 2011 Jun 1;3(4):1216-31. doi: 10.2741/222. Front Biosci (Schol Ed). 2011. PMID: 21622267 Review.
Cited by
-
Regulation of Tight Junctions by Sex Hormones in Goat Mammary Epithelial Cells.Animals (Basel). 2022 May 30;12(11):1404. doi: 10.3390/ani12111404. Animals (Basel). 2022. PMID: 35681868 Free PMC article.
-
Pathophysiological Changes and the Role of Notch-1 Activation After Decompression in a Compressive Spinal Cord Injury Rat Model.Front Neurosci. 2021 Jan 28;15:579431. doi: 10.3389/fnins.2021.579431. eCollection 2021. Front Neurosci. 2021. PMID: 33584186 Free PMC article.
-
Understanding the Blood-Brain Barrier: From Physiology to Pathology.Adv Exp Med Biol. 2025;1477:1-33. doi: 10.1007/978-3-031-89525-8_1. Adv Exp Med Biol. 2025. PMID: 40442381 Review.
-
Sex-Related Differences of Matrix Metalloproteinases (MMPs): New Perspectives for These Biomarkers in Cardiovascular and Neurological Diseases.J Pers Med. 2022 Jul 22;12(8):1196. doi: 10.3390/jpm12081196. J Pers Med. 2022. PMID: 35893290 Free PMC article. Review.
-
Nuclear receptor ligand screening in an iPSC-derived in vitro blood-brain barrier model identifies new contributors to leptin transport.Fluids Barriers CNS. 2022 Sep 21;19(1):77. doi: 10.1186/s12987-022-00375-3. Fluids Barriers CNS. 2022. PMID: 36131285 Free PMC article.
References
-
- Hawkins BT, Davis TP. The blood-brain barrier/neurovascular unit in health and disease. Pharmacol Rev. 2005;57:173–185. - PubMed
-
- Bartanusz V, Jezova D, Alajajian B, Digicaylioglu M. The blood-spinal cord barrier: morphology and clinical implications. Ann Neurol. 2011;70:194–206. - PubMed
-
- Haseloff RF, Dithmer S, Winkler L, Wolburg H, Blasig IE. Transmembrane proteins of the tight junctions at the blood-brain barrier: Structural and functional aspects. Semin Cell Dev Biol. 2014;38:16–25. - PubMed
-
- Drewes LR. Molecular architecture of the brain microvasculature: perspective on blood-brain transport. J Mol Neurosci. 2001;16:93–98. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
