

Interacting Effects of Light and Iron Availability on the Coupling of Photosynthetic Electron Transport and CO2-Assimilation in Marine Phytoplankton
- PMID: 26171963
- PMCID: PMC4501554
- DOI: 10.1371/journal.pone.0133235
Interacting Effects of Light and Iron Availability on the Coupling of Photosynthetic Electron Transport and CO2-Assimilation in Marine Phytoplankton
Abstract
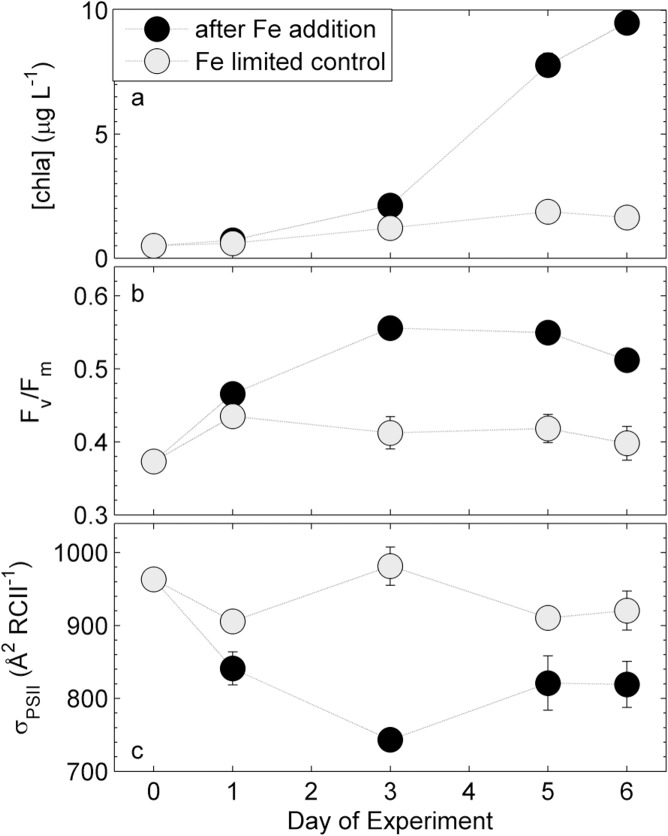
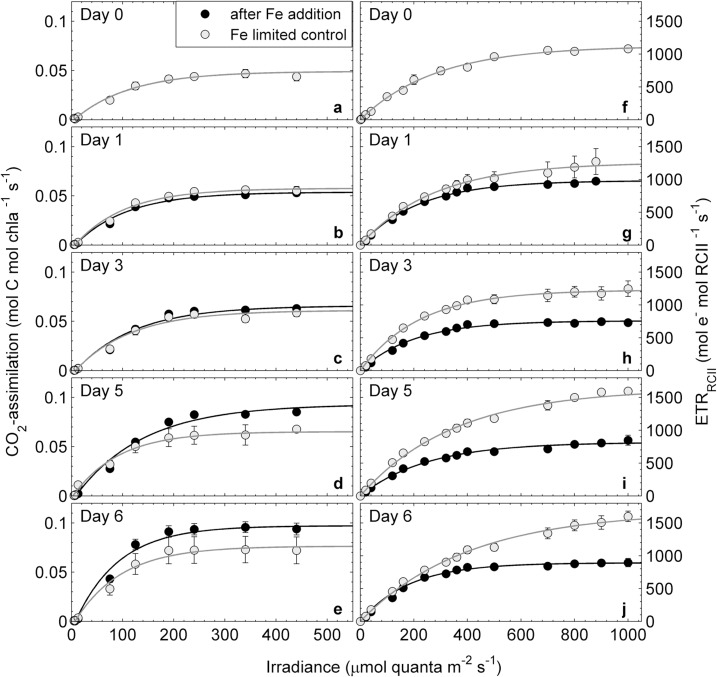
Iron availability directly affects photosynthesis and limits phytoplankton growth over vast oceanic regions. For this reason, the availability of iron is a crucial variable to consider in the development of active chlorophyll a fluorescence based estimates of phytoplankton primary productivity. These bio-optical approaches require a conversion factor to derive ecologically-relevant rates of CO2-assimilation from estimates of electron transport in photosystem II. The required conversion factor varies significantly across phytoplankton taxa and environmental conditions, but little information is available on its response to iron limitation. In this study, we examine the role of iron limitation, and the interacting effects of iron and light availability, on the coupling of photosynthetic electron transport and CO2-assimilation in marine phytoplankton. Our results show that excess irradiance causes increased decoupling of carbon fixation and electron transport, particularly under iron limiting conditions. We observed that reaction center II specific rates of electron transport (ETR(RCII), mol e- mol RCII(-1) s(-1)) increased under iron limitation, and we propose a simple conceptual model for this observation. We also observed a strong correlation between the derived conversion factor and the expression of non-photochemical quenching. Utilizing a dataset from in situ phytoplankton assemblages across a coastal--oceanic transect in the Northeast subarctic Pacific, this relationship was used to predict ETR(RCII): CO2-assimilation conversion factors and carbon-based primary productivity from FRRF data, without the need for any additional measurements.
Conflict of interest statement
Figures







Similar articles
-
Phytoplankton Productivity in an Arctic Fjord (West Greenland): Estimating Electron Requirements for Carbon Fixation and Oxygen Production.PLoS One. 2015 Jul 28;10(7):e0133275. doi: 10.1371/journal.pone.0133275. eCollection 2015. PLoS One. 2015. PMID: 26218096 Free PMC article.
-
Development of photosynthetic carbon fixation model using multi-excitation wavelength fast repetition rate fluorometry in Lake Biwa.PLoS One. 2021 Feb 2;16(2):e0238013. doi: 10.1371/journal.pone.0238013. eCollection 2021. PLoS One. 2021. PMID: 33529253 Free PMC article.
-
Comparison by PAM fluorometry of photosynthetic activity of nine marine phytoplankton grown under identical conditions.Photochem Photobiol. 2005 May-Jun;81(3):649-53. doi: 10.1562/2005-01-13-RA-414. Photochem Photobiol. 2005. PMID: 15686444
-
Iron, phytoplankton growth, and the carbon cycle.Met Ions Biol Syst. 2005;43:153-93. doi: 10.1201/9780824751999.ch7. Met Ions Biol Syst. 2005. PMID: 16370118 Review.
-
Computer modeling of electron and proton transport in chloroplasts.Biosystems. 2014 Jul;121:1-21. doi: 10.1016/j.biosystems.2014.04.007. Epub 2014 May 14. Biosystems. 2014. PMID: 24835748 Review.
Cited by
-
Fast Repetition Rate Fluorometry (FRRF) Derived Phytoplankton Primary Productivity in the Bay of Bengal.Front Microbiol. 2019 May 24;10:1164. doi: 10.3389/fmicb.2019.01164. eCollection 2019. Front Microbiol. 2019. PMID: 31244786 Free PMC article.
-
Impact of Lhcx2 on Acclimation to Low Iron Conditions in the Diatom Phaeodactylum tricornutum.Front Plant Sci. 2022 Mar 16;13:841058. doi: 10.3389/fpls.2022.841058. eCollection 2022. Front Plant Sci. 2022. PMID: 35371185 Free PMC article.
-
Iron and manganese co-limit growth of the Southern Ocean diatom Chaetoceros debilis.PLoS One. 2019 Sep 16;14(9):e0221959. doi: 10.1371/journal.pone.0221959. eCollection 2019. PLoS One. 2019. PMID: 31525212 Free PMC article.
-
Irradiance and nutrient-dependent effects on photosynthetic electron transport in Arctic phytoplankton: A comparison of two chlorophyll fluorescence-based approaches to derive primary photochemistry.PLoS One. 2021 Dec 9;16(12):e0256410. doi: 10.1371/journal.pone.0256410. eCollection 2021. PLoS One. 2021. PMID: 34882695 Free PMC article.
-
Resistance of Arctic phytoplankton to ocean acidification and enhanced irradiance.Polar Biol. 2018;41(3):399-413. doi: 10.1007/s00300-017-2186-0. Epub 2017 Aug 9. Polar Biol. 2018. PMID: 31983801 Free PMC article.
References
-
- Field CB, Behrenfeld MJ, Randerson JT, Falkowski P. Primary production of the biosphere: integrating terrestrial and oceanic components. Science. 1998;281: 237–240. - PubMed
-
- Beardall J, Raven JA. The potential effects of global climate change on microalgal photosynthesis, growth and ecology. Phycologia. 2004;43: 26–40. 10.2216/i0031-8884-43-1-26.1 - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
