

Review
doi: 10.1038/nrn3921.
Epub 2015 Jul 15.
Development and functions of the choroid plexus-cerebrospinal fluid system
Affiliations
- PMID: 26174708
- PMCID: PMC4629451
- DOI: 10.1038/nrn3921
Item in Clipboard
Review
Development and functions of the choroid plexus-cerebrospinal fluid system
Nat Rev Neurosci.
2015 Aug.
Abstract
The choroid plexus (ChP) is the principal source of cerebrospinal fluid (CSF), which has accepted roles as a fluid cushion and a sink for nervous system waste in vertebrates. Various animal models have provided insights into how the ChP-CSF system develops and matures. In addition, recent studies have uncovered new, active roles for this dynamic system in the regulation of neural stem cells, critical periods and the overall health of the nervous system. Together, these findings have brought about a paradigm shift in our understanding of brain development and health, and have stimulated new initiatives for the treatment of neurological disease.
Figures
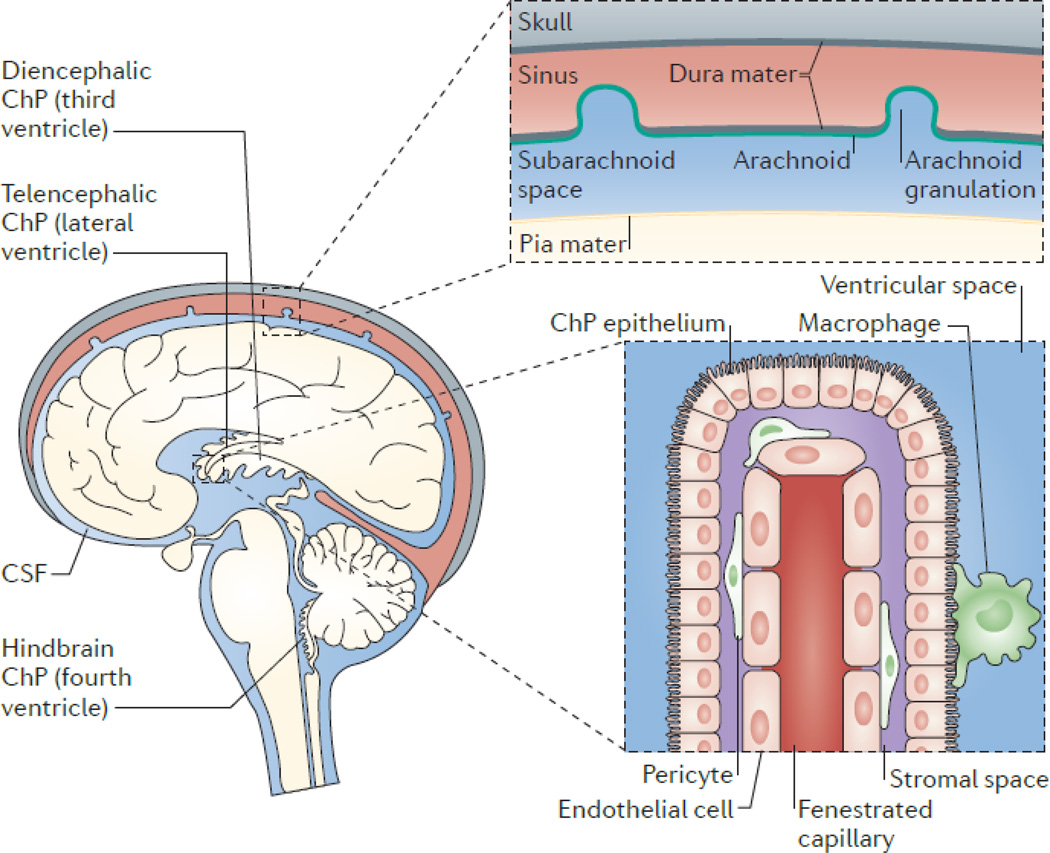
The choroid plexus (ChP) consists of epithelial cells that surround a core of capillaries and connective tissue. The epithelial cells are joined by tight junctions, which form the blood–cerebrospinal fluid (CSF) barrier. The ChP is located in each ventricle in the brain, and is regarded as the principal source of CSF, secreting up to 500ml of CSF per day in the adult human brain. CSF flows from the lateral to the third ventricle through the interventricular foramina, and from the third to the fourth ventricle through the cerebral aqueduct. CSF can then enter the central canal of the spinal cord or the subarachnoid space, where microscopic arachnoid villi and macroscopic arachnoid granulations (granulations are present in humans and other large mammals), together with lymphatics present in the cribriform plate–olfactory region– or lining the meninges-dural sinuses (6 - Louveau), resorb CSF into the systemic circulation or into regional and cervical lymph nodes. The ChP is organized into an outer layer of cuboidal epithelial cells surrounding a core of fenestrated capillaries and other stromal cell types. Functioning as the blood-CSF barrier, the ChP is also a gateway for immune cell entry into the central nervous system (BOX 1).
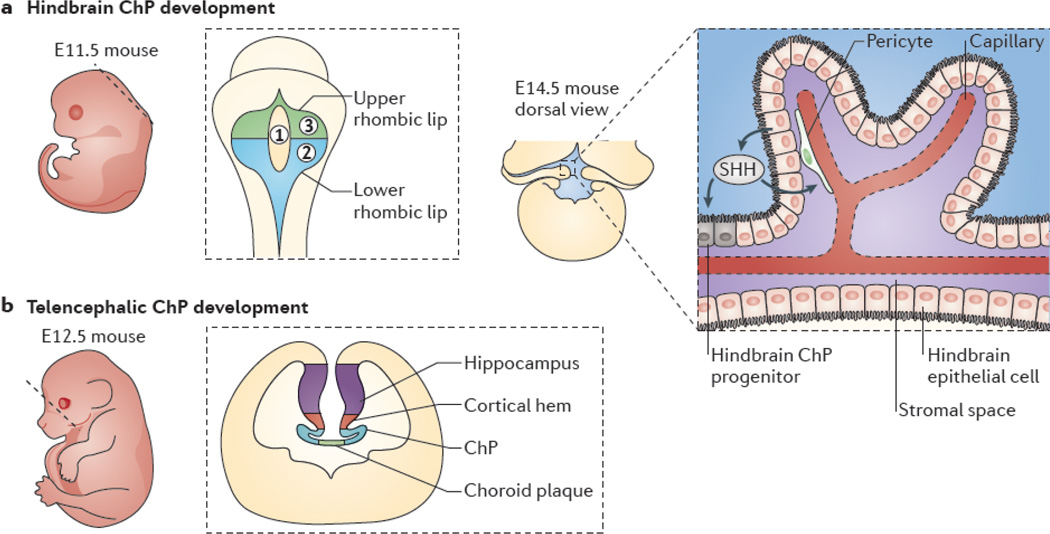
a | Hindbrain choroid plexus (ChP) development begins at about embryonic day 9.5 (E9.5) and peaks between ∼E11 and E12. The left panel shows a sagittal view of mouse embryo at E11.5; the dashed line represents idealized field of view for the middle panel, which shows a dorsal view of the neural tube. The rhombic lip neuroepithelium (gray) surrounds three fields of hindbrain roofplate epithelium. Field 1 (yellow) contains cells that do not express transthyretin (Ttr) or potassium voltage-gated channel subfamily E member 2 (Kcne2), whereas field 2 (blue) contains caudally derived (from rhombomeres 2–8) Ttr- and Kcne2-positive cells and field 3 (green) contains rostrally derived (from rhombomere 1) Ttr- and Kcne2-positive cells. The right panel depicts the dorsal view of a proliferating ChP, as it emerges from the lateral edges of the fourth ventricle and grows medially to complete its development by E14. Sonic hedgehog (SHH) that is secreted by hindbrain ChP epithelial cells (black) signals to underlying pericytes (green) to regulate vascular outgrowth (red) while also signalling to hindbrain ChP progenitors adjacent to the lower rhombic lip to induce further progenitor proliferation (inset). b | Development of telencephalic ChP follows that of the hindbrain ChP. The left panel shows a sagittal view of a mouse embryo at E12.5, in which the dashed line represents the idealized coronal section that is shown in the right panel. The right panel depicts a telencephalic ChP developing bilaterally at the invaginated dorsal midline of the neural tube. Telencephalic ChP formation requires signals from the dorsal midline, including bone morphogenetic proteins and WNTs originating from the cortical hem. Disruption of the dorsal midline is frequently associated with either ChP over proliferation or failure of the ChP to form.
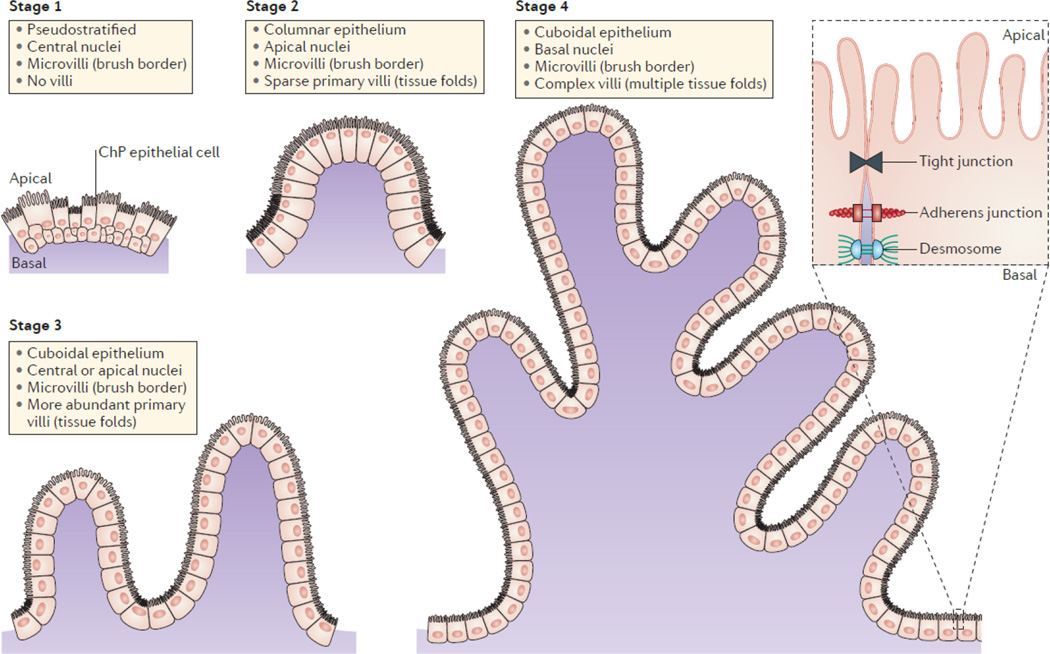
The choroid plexus (ChP) of each ventricle has the same morphology and is organized into a layer of cuboidal epithelium surrounding a core of vasculature and other stromal cells. The maturation of epithelial cells includes four stereotypical stages. The ChP epithelial cells first appear to be pseudostratified with centrally located nuclei in stage 1 with little to no villous elaboration. This is followed by a transition to a columnar epithelium with apically located nuclei in stage 2, and the tissue becomes convoluted into sparse primary villi. In stage 3, the epithelial cells flatten to become more cuboidal in shape, nuclei are centrally or apically located, and primary villi in the tissue become more abundant. Last, in stage 4, nuclei become more basally located, which is thought to be caused by the apical enrichment of transport machinery and microvilli, and villi are more complex with multiple fronds. Junctions and adhesion molecules are found between adjacent cells, allowing the mature ChP to function as the blood–cerebrospinal fluid (CSF) barrier by restricting the passage of solutes from systemic circulation into CSF.
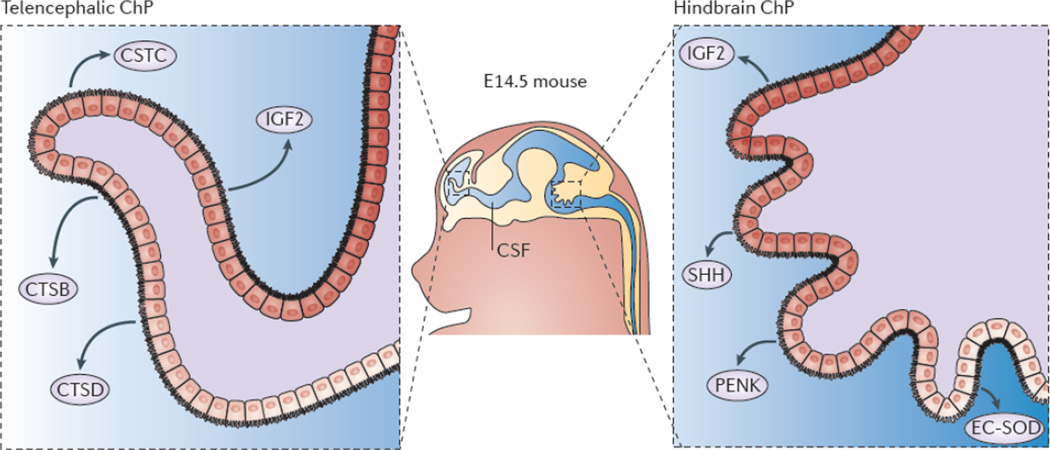
Gene expression profiling of mouse and primate telencephalic choroid plexus (ChP) and hindbrain ChP reveal that, despite being morphologically similar, these tissues are transcriptionally heterogeneous. The ChPs maintain distinct positional identities reminiscent of their axial tissues of origin. The telencephalic ChP has higher expression levels of Emx2Otx1, and Six3, which are markers of the telencephalon, whereas the hindbrain ChP has higher expression levels of Hoxa2En2, and Meis1, genes critical for patterning the hindbrain. Further analysis of the ChP secretome reveals that these tissues are functionally distinct in their expression and secretion of signalling factors into the cerebrospinal fluid (CSF). In particular, the telencephalic ChP secretes more cystatin C (CSTC), cathepsin B (CTSB) and cathepsin D (CTSD), whereas the hindbrain ChP secretes more Sonic hedgehog (SHH), proenkephalin (PENK) and extracellular superoxide-dismutase (EC-SOD). Protein expression domains have been observed within each ChP (for example, EC-SOD and SHH protein domains), which is suggestive of protein gradients within each ventricle. These findings indicate that the molecular heterogeneity of the telencephalic versus the hindbrain ChP contributes to establishing a regionalized CSF and support a model of protein gradients throughout the ventricular system.
References
-
- Damkier HH, Brown PD, Praetorius J. Cerebrospinal fluid secretion by the choroid plexus. Physiol Rev. 2013;93:1847–1892. - PubMed
-
- Koh L, et al. Development of cerebrospinal fluid absorption sites in the pig and rat: connections between the subarachnoid space and lymphatic vessels in the olfactory turbinates. Anat Embryol (Berl) 2006;211:335–344. - PubMed
-
- Johnston M, Zakharov A, Koh L, Armstrong D. Subarachnoid injection of Microfil reveals connections between cerebrospinal fluid and nasal lymphatics in the non-human primate. Neuropathol Appl Neurobiol. 2005;31:632–640. - PubMed
-
- Mollanji R, Bozanovic-Sosic R, Zakharov A, Makarian L, Johnston MG. Blocking cerebrospinal fluid absorption through the cribriform plate increases resting intracranial pressure. Am J Physiol Regul Integr Comp Physiol. 2002;282:R1593–R1599. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
