

Emerging and Novel Functions of Complement Protein C1q
- PMID: 26175731
- PMCID: PMC4484229
- DOI: 10.3389/fimmu.2015.00317
Emerging and Novel Functions of Complement Protein C1q
Abstract
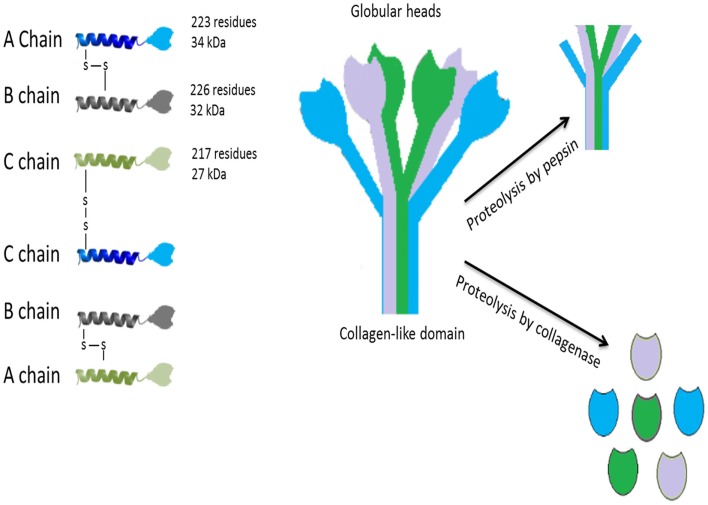
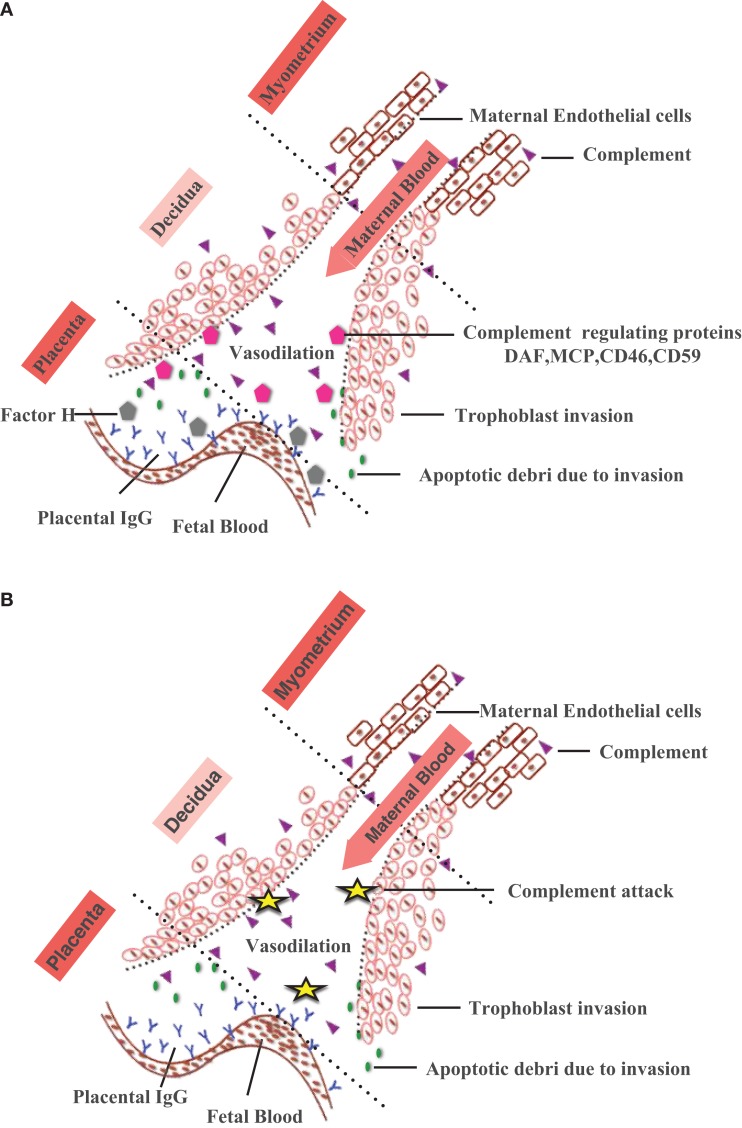
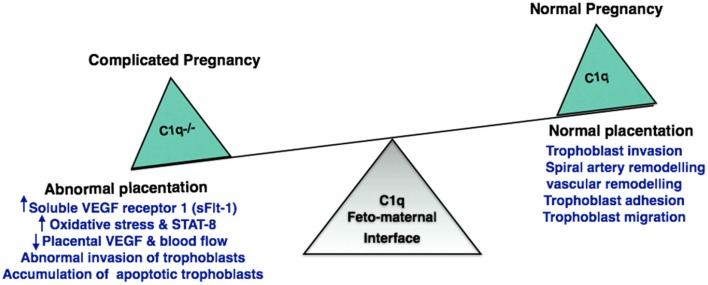
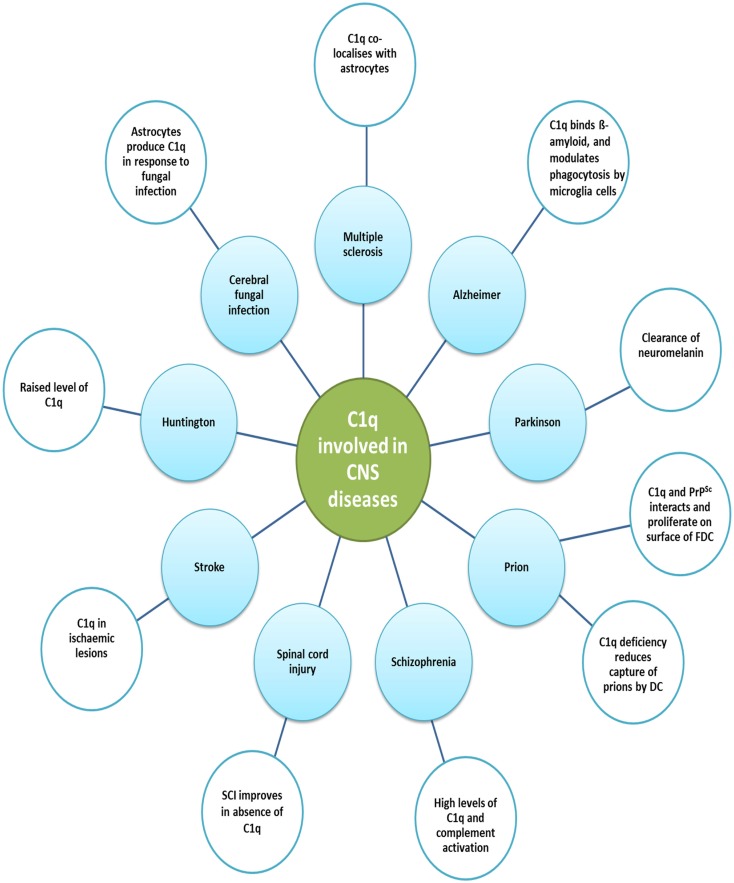
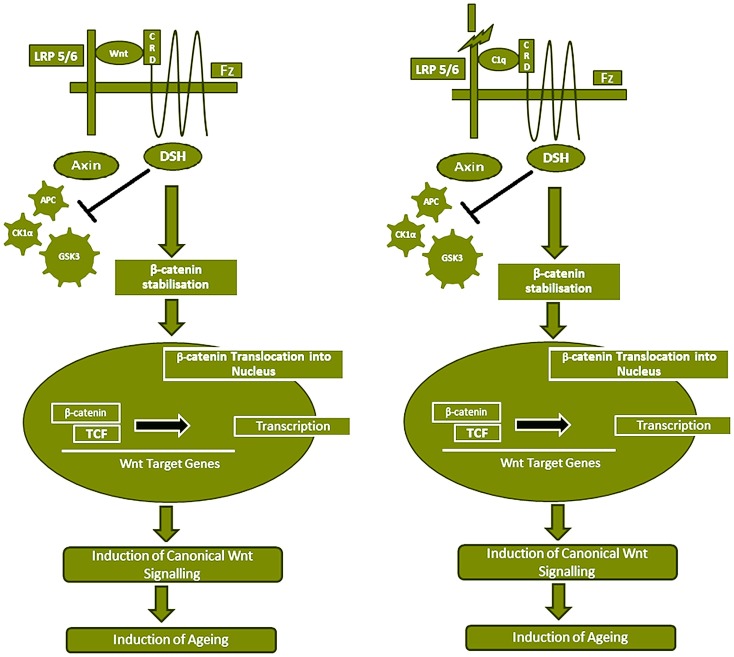
Complement protein C1q, the recognition molecule of the classical pathway, performs a diverse range of complement and non-complement functions. It can bind various ligands derived from self, non-self, and altered self and modulate the functions of immune and non-immune cells including dendritic cells and microglia. C1q involvement in the clearance of apoptotic cells and subsequent B cell tolerance is more established now. Recent evidence appears to suggest that C1q plays an important role in pregnancy where its deficiency and dysregulation can have adverse effects, leading to preeclampsia, missed abortion, miscarriage or spontaneous loss, and various infections. C1q is also produced locally in the central nervous system, and has a protective role against pathogens and possible inflammatory functions while interacting with aggregated proteins leading to neurodegenerative diseases. C1q role in synaptic pruning, and thus CNS development, its anti-cancer effects as an immune surveillance molecule, and possibly in aging are currently areas of extensive research.
Keywords: C1q; apoptosis; cancer; complement; neurogenesis; pregnancy.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources