

The SMC Loader Scc2 Promotes ncRNA Biogenesis and Translational Fidelity
- PMID: 26176819
- PMCID: PMC4503661
- DOI: 10.1371/journal.pgen.1005308
The SMC Loader Scc2 Promotes ncRNA Biogenesis and Translational Fidelity
Abstract
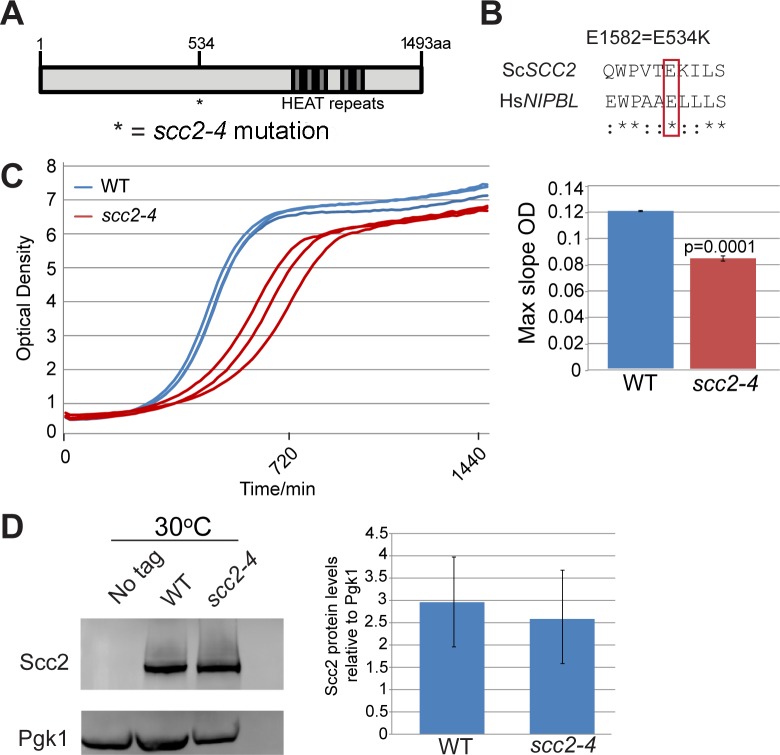
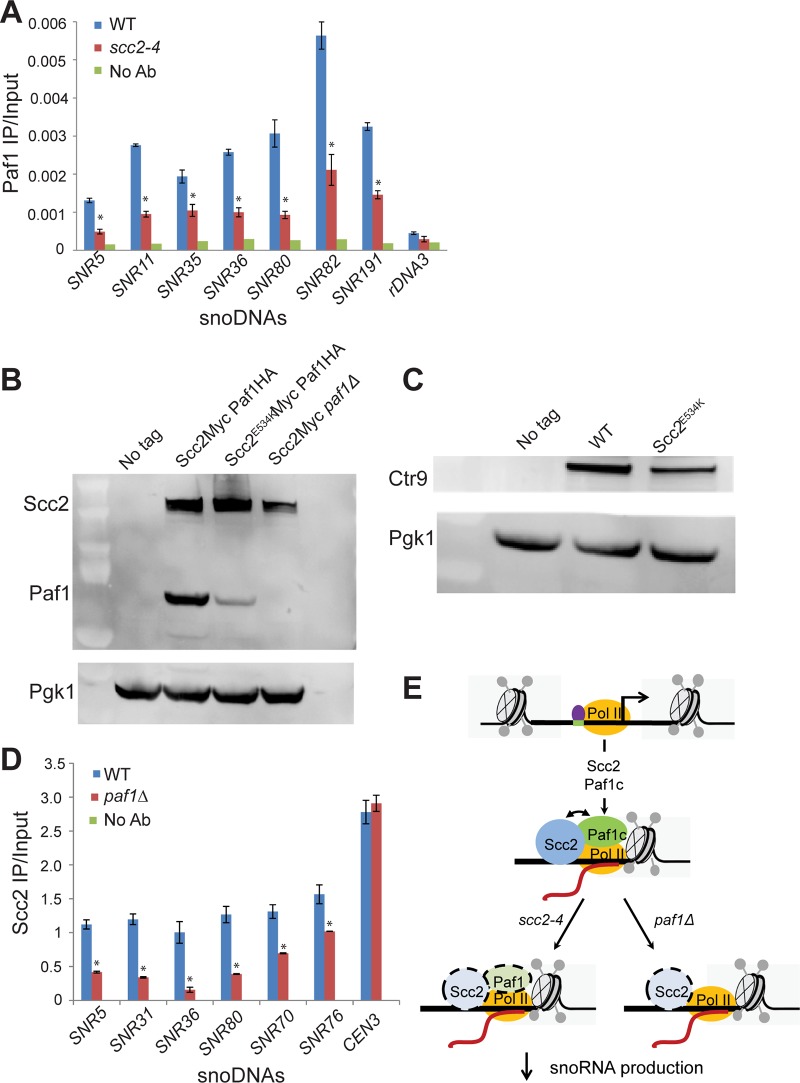
The Scc2-Scc4 complex is essential for loading the cohesin complex onto DNA. Cohesin has important roles in chromosome segregation, DSB repair, and chromosome condensation. Here we report that Scc2 is important for gene expression in budding yeast. Scc2 and the transcriptional regulator Paf1 collaborate to promote the production of Box H/ACA snoRNAs which guide pseudouridylation of RNAs including ribosomal RNA. Mutation of SCC2 was associated with defects in the production of ribosomal RNA, ribosome assembly, and splicing. While the scc2 mutant does not have a general defect in protein synthesis, it shows increased frameshifting and reduced cap-independent translation. These findings suggest Scc2 normally promotes a gene expression program that supports translational fidelity. We hypothesize that translational dysfunction may contribute to the human disorder Cornelia de Lange syndrome, which is caused by mutations in NIPBL, the human ortholog of SCC2.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






Similar articles
-
Mediator recruits the cohesin loader Scc2 to RNA Pol II-transcribed genes and promotes sister chromatid cohesion.Curr Biol. 2022 Jul 11;32(13):2884-2896.e6. doi: 10.1016/j.cub.2022.05.019. Epub 2022 Jun 1. Curr Biol. 2022. PMID: 35654035 Free PMC article.
-
New insights into cohesin loading.Curr Genet. 2018 Feb;64(1):53-61. doi: 10.1007/s00294-017-0723-6. Epub 2017 Jun 19. Curr Genet. 2018. PMID: 28631016 Review.
-
The Scc2-Scc4 complex acts in sister chromatid cohesion and transcriptional regulation by maintaining nucleosome-free regions.Nat Genet. 2014 Oct;46(10):1147-51. doi: 10.1038/ng.3080. Epub 2014 Aug 31. Nat Genet. 2014. PMID: 25173104 Free PMC article.
-
Cohesin proteins promote ribosomal RNA production and protein translation in yeast and human cells.PLoS Genet. 2012;8(6):e1002749. doi: 10.1371/journal.pgen.1002749. Epub 2012 Jun 14. PLoS Genet. 2012. PMID: 22719263 Free PMC article.
-
NIPBL and cohesin: new take on a classic tale.Trends Cell Biol. 2023 Oct;33(10):860-871. doi: 10.1016/j.tcb.2023.03.006. Epub 2023 Apr 15. Trends Cell Biol. 2023. PMID: 37062615 Review.
Cited by
-
E3 ubiquitin ligase Bre1 couples sister chromatid cohesion establishment to DNA replication in Saccharomyces cerevisiae.Elife. 2017 Oct 23;6:e28231. doi: 10.7554/eLife.28231. Elife. 2017. PMID: 29058668 Free PMC article.
-
Structure of the cohesin loader Scc2.Nat Commun. 2017 Jan 6;8:13952. doi: 10.1038/ncomms13952. Nat Commun. 2017. PMID: 28059076 Free PMC article.
-
NIPBL Controls RNA Biogenesis to Prevent Activation of the Stress Kinase PKR.Cell Rep. 2016 Jan 5;14(1):93-102. doi: 10.1016/j.celrep.2015.12.012. Epub 2015 Dec 24. Cell Rep. 2016. PMID: 26725122 Free PMC article.
-
Phosphorylation of the Scc2 cohesin deposition complex subunit regulates chromosome condensation through cohesin integrity.Mol Biol Cell. 2015 Nov 1;26(21):3754-67. doi: 10.1091/mbc.E15-03-0165. Epub 2015 Sep 9. Mol Biol Cell. 2015. PMID: 26354421 Free PMC article.
-
Chl1 DNA helicase and Scc2 function in chromosome condensation through cohesin deposition.PLoS One. 2017 Nov 29;12(11):e0188739. doi: 10.1371/journal.pone.0188739. eCollection 2017. PLoS One. 2017. PMID: 29186203 Free PMC article.
References
-
- Michaelis C, Ciosk R, Nasmyth K. Cohesins: chromosomal proteins that prevent premature separation of sister chromatids. Cell. 1997. October 3;91(1):35–45. - PubMed
-
- Gruber S, Haering CH, Nasmyth K. Chromosomal cohesin forms a ring. Cell. 2003. March 21;112(6):765–77. - PubMed
-
- Haering CH, Lowe J, Hochwagen A, Nasmyth K. Molecular architecture of SMC proteins and the yeast cohesin complex. Molecular cell. 2002. April;9(4):773–88. - PubMed
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
