

The virus-induced protein APOBEC3G inhibits anoikis by activation of Akt kinase in pancreatic cancer cells
- PMID: 26178819
- PMCID: PMC4503957
- DOI: 10.1038/srep12230
The virus-induced protein APOBEC3G inhibits anoikis by activation of Akt kinase in pancreatic cancer cells
Abstract
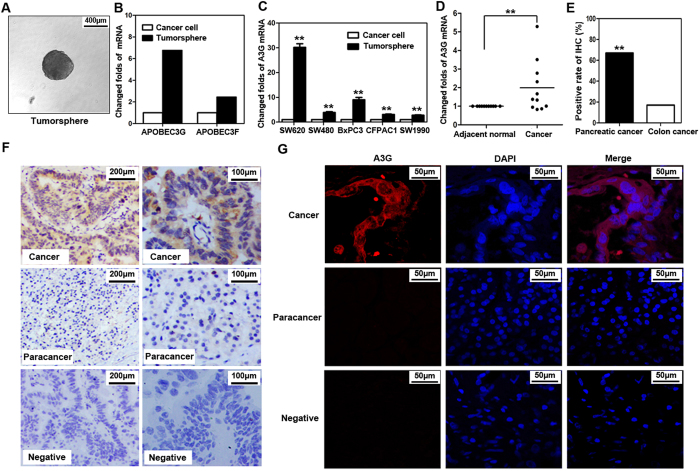
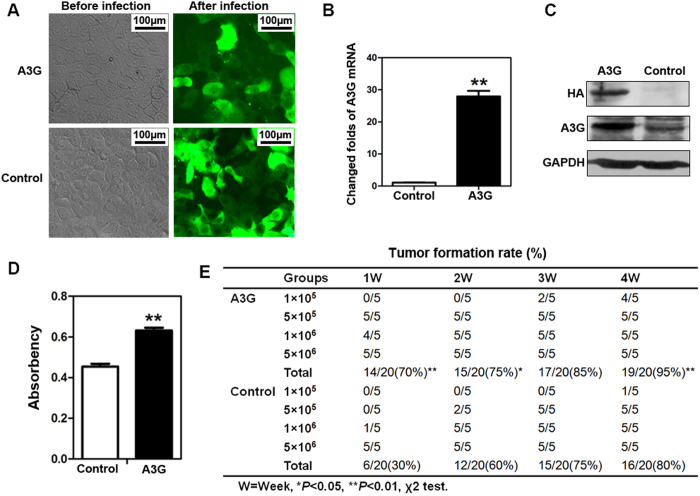
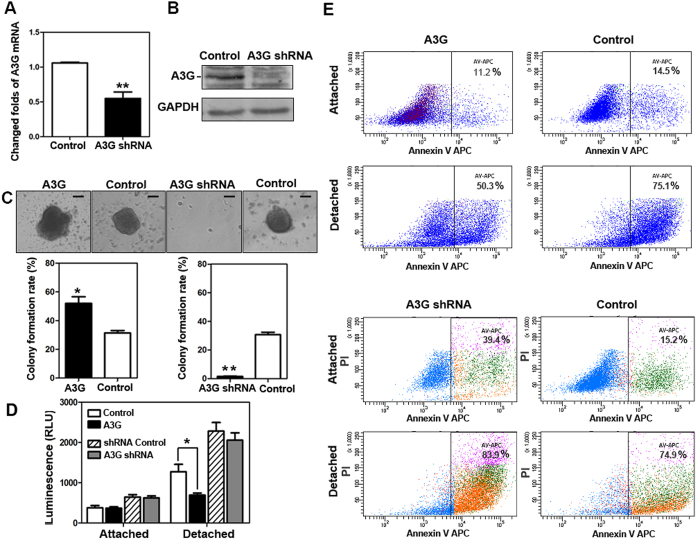
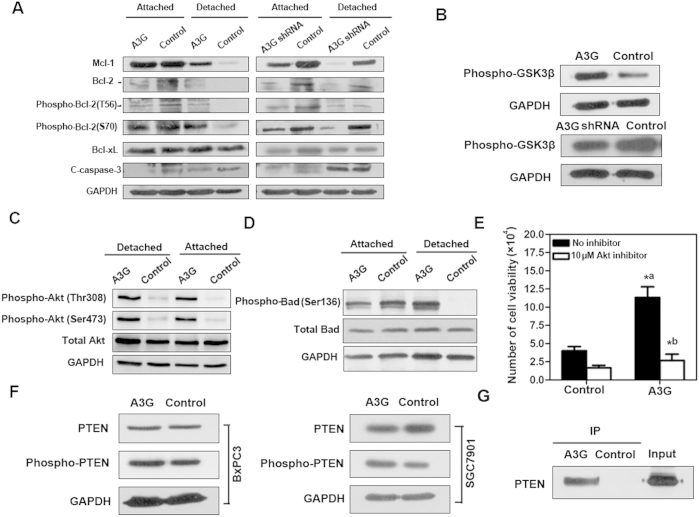
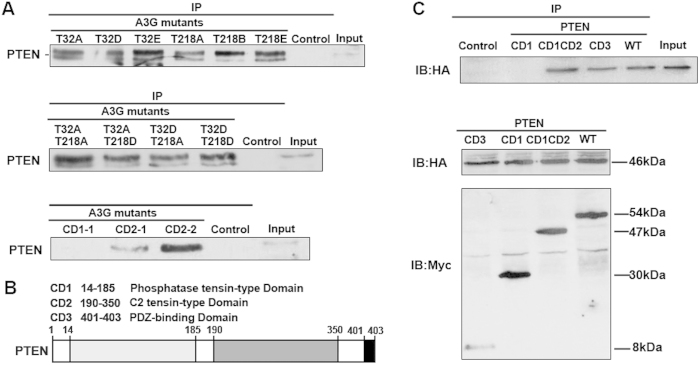
Pancreatic cancer is one of the more common cancers with a poor prognosis. Some varieties of cancer are related to virus infection. As a virus-induced protein, APOBEC3G (A3G) presents extensive anti-virus ability, but the role of A3G in pancreatic cancer was previously unknown. The expression of A3G in pancreatic cancer was examined using TaqMan real-time qPCR, immunohistochemical and immunofluorescent staining. Subsequently, the role of A3G in pancreatic cancer was evaluated in vivo using the tumor xenograft model. Anoikis was detected by colony formation assay and flow cytometry in vitro. The Akt kinase activity and target protein PTEN were examined by co-immunoprecipitation and immunoblot. The virus-induced protein A3G was significantly up-regulated in pancreatic cancer, and the up-regulation of A3G promoted xenograft tumor formation. A3G inactivated PTEN by binding to the C2 tensin-type and PDZ domains, thereby inducing anoikis resistance through Akt activation. Our results demonstrate that the up-regulation of A3G in pancreatic cancer cells induces anoikis resistance, and they provide novel insight into the mechanism by which A3G affects the malignant behavior of pancreatic cancer cells.
Figures
Similar articles
-
Tissue transglutaminase regulates focal adhesion kinase/AKT activation by modulating PTEN expression in pancreatic cancer cells.Clin Cancer Res. 2008 Apr 1;14(7):1997-2005. doi: 10.1158/1078-0432.CCR-07-1533. Clin Cancer Res. 2008. PMID: 18381937
-
Hepatocyte growth factor inhibits anoikis of pancreatic carcinoma cells through phosphatidylinositol 3-kinase pathway.Pancreas. 2011 May;40(4):608-14. doi: 10.1097/MPA.0b013e318214fa6c. Pancreas. 2011. PMID: 21499215
-
Overexpression of HMGA1 promotes anoikis resistance and constitutive Akt activation in pancreatic adenocarcinoma cells.Br J Cancer. 2007 Mar 26;96(6):993-1000. doi: 10.1038/sj.bjc.6603654. Epub 2007 Mar 6. Br J Cancer. 2007. Retraction in: Br J Cancer. 2025 Jun;132(12):1200. doi: 10.1038/s41416-025-03036-y. PMID: 17342093 Free PMC article. Retracted.
-
Regulated production and anti-HIV type 1 activities of cytidine deaminases APOBEC3B, 3F, and 3G.AIDS Res Hum Retroviruses. 2005 Jul;21(7):611-9. doi: 10.1089/aid.2005.21.611. AIDS Res Hum Retroviruses. 2005. PMID: 16060832
-
Anoikis resistance and oncoviruses.J Cell Biochem. 2018 Mar;119(3):2484-2491. doi: 10.1002/jcb.26363. Epub 2017 Sep 12. J Cell Biochem. 2018. PMID: 28836703 Review.
Cited by
-
Next generation APOBEC3 inhibitors: optimally designed for potency and nuclease stability.Nucleic Acids Res. 2025 Mar 20;53(6):gkaf234. doi: 10.1093/nar/gkaf234. Nucleic Acids Res. 2025. PMID: 40156866 Free PMC article.
-
A Conserved uORF Regulates APOBEC3G Translation and Is Targeted by HIV-1 Vif Protein to Repress the Antiviral Factor.Biomedicines. 2021 Dec 22;10(1):13. doi: 10.3390/biomedicines10010013. Biomedicines. 2021. PMID: 35052693 Free PMC article.
-
Next generation APOBEC3 inhibitors: Optimally designed for potency and nuclease stability.bioRxiv [Preprint]. 2024 Sep 6:2024.09.05.611238. doi: 10.1101/2024.09.05.611238. bioRxiv. 2024. Update in: Nucleic Acids Res. 2025 Mar 20;53(6):gkaf234. doi: 10.1093/nar/gkaf234. PMID: 39282427 Free PMC article. Updated. Preprint.
-
Barriers to Infection of Human Cells by Feline Leukemia Virus: Insights into Resistance to Zoonosis.J Virol. 2017 Feb 14;91(5):e02119-16. doi: 10.1128/JVI.02119-16. Print 2017 Mar 1. J Virol. 2017. PMID: 28031367 Free PMC article.
-
UHMK1 Is a Novel Marker for Personalized Prediction of Pancreatic Cancer Prognosis.Front Oncol. 2022 Mar 10;12:834647. doi: 10.3389/fonc.2022.834647. eCollection 2022. Front Oncol. 2022. PMID: 35359403 Free PMC article.
References
-
- Siegel R., Naishadham D. & Jemal A. Cancer statistics, 2012. CA Cancer J Clin. 62, 10–29 (2012). - PubMed
-
- Michl P. & Gress T. M. Current concepts and novel targets in advanced pancreatic cancer. Gut. 60, 317–326 (2013). - PubMed
-
- Douma S. et al. Suppression of anoikis and induction of metastasis by the neurotrophic receptor TrkB. Nature. 430, 1034–1039 (2004). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
