

Rho1- and Pkc1-dependent phosphorylation of the F-BAR protein Syp1 contributes to septin ring assembly
- PMID: 26179915
- PMCID: PMC4569315
- DOI: 10.1091/mbc.E15-06-0366
Rho1- and Pkc1-dependent phosphorylation of the F-BAR protein Syp1 contributes to septin ring assembly
Abstract
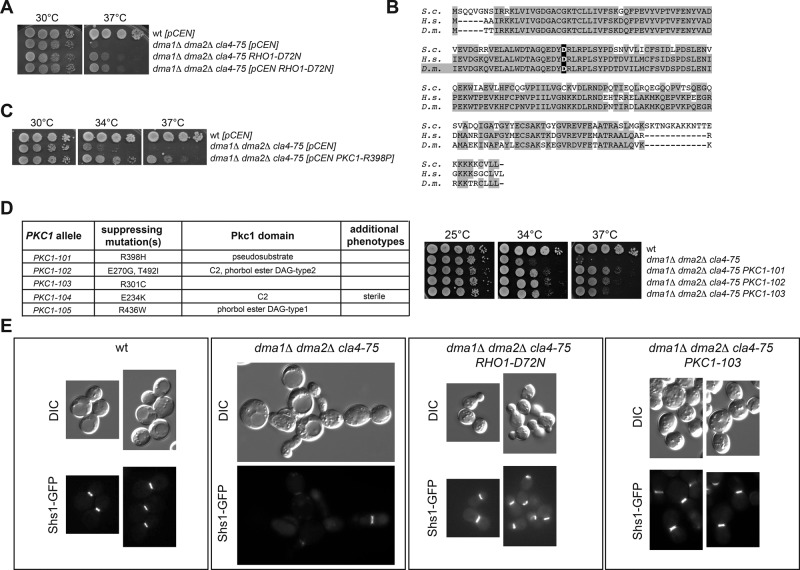
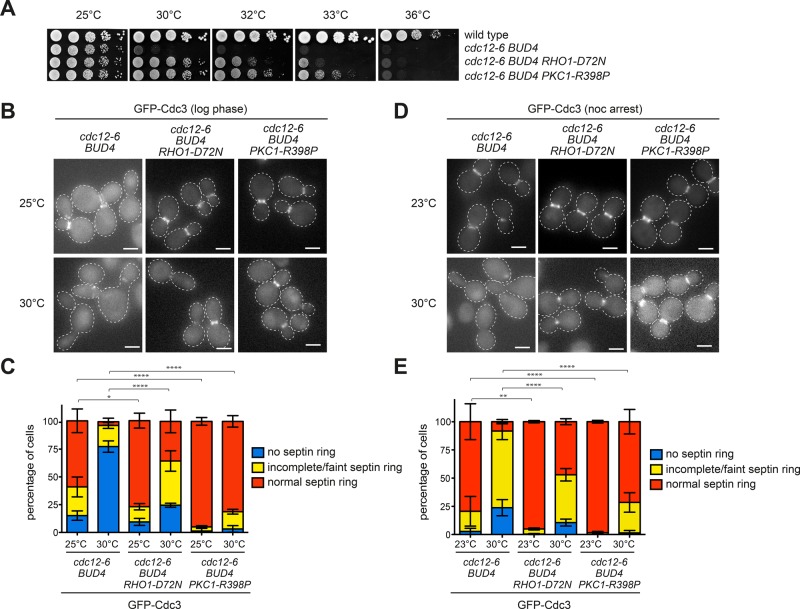
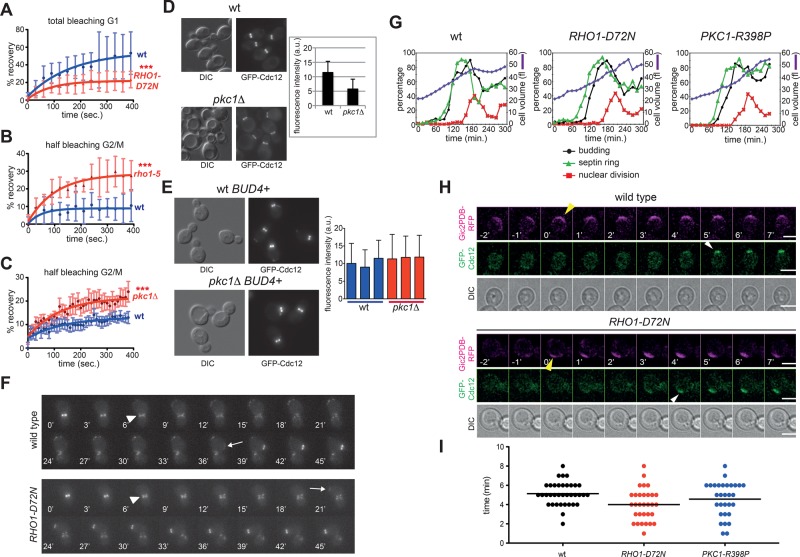
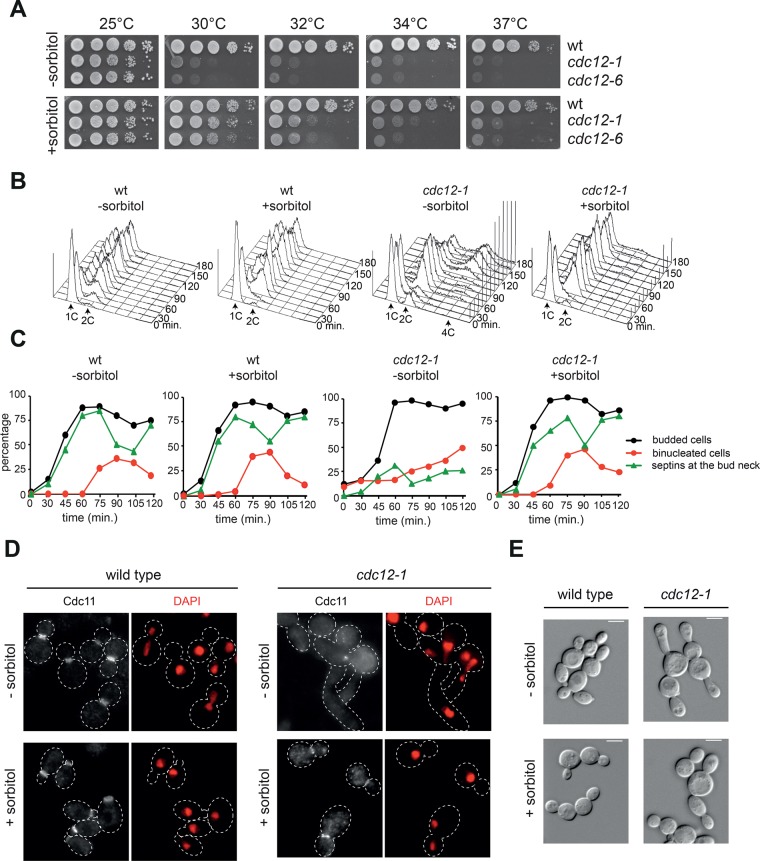
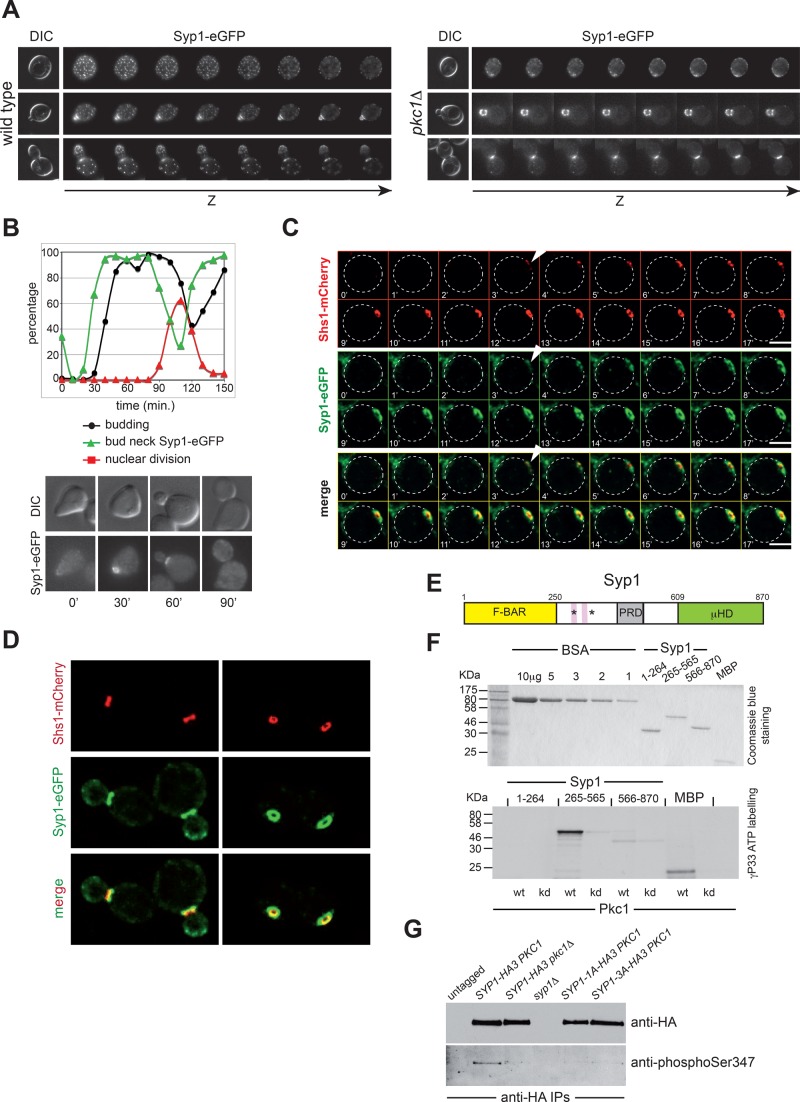
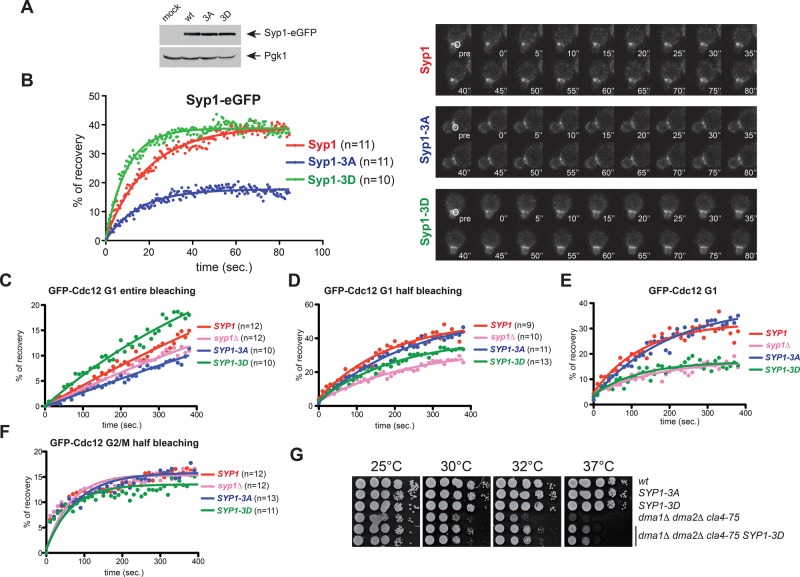
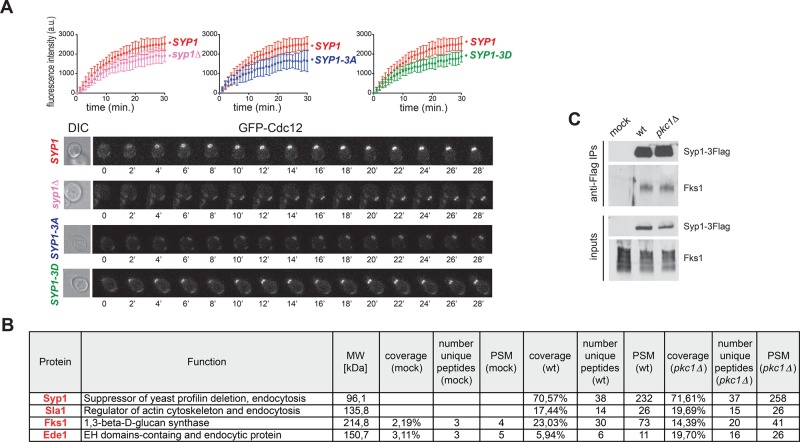
In many cell types, septins assemble into filaments and rings at the neck of cellular appendages and/or at the cleavage furrow to help compartmentalize the plasma membrane and support cytokinesis. How septin ring assembly is coordinated with membrane remodeling and controlled by mechanical stress at these sites is unclear. Through a genetic screen, we uncovered an unanticipated link between the conserved Rho1 GTPase and its effector protein kinase C (Pkc1) with septin ring stability in yeast. Both Rho1 and Pkc1 stabilize the septin ring, at least partly through phosphorylation of the membrane-associated F-BAR protein Syp1, which colocalizes asymmetrically with the septin ring at the bud neck. Syp1 is displaced from the bud neck upon Pkc1-dependent phosphorylation at two serines, thereby affecting the rigidity of the new-forming septin ring. We propose that Rho1 and Pkc1 coordinate septin ring assembly with membrane and cell wall remodeling partly by controlling Syp1 residence at the bud neck.
© 2015 Merlini et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
Similar articles
-
Sequential recruitment of F-BAR proteins controls cytoskeletal crosstalk at the yeast bud neck.Curr Biol. 2025 Feb 3;35(3):574-590.e10. doi: 10.1016/j.cub.2024.12.011. Epub 2025 Jan 10. Curr Biol. 2025. PMID: 39798561
-
Cell cycle control of septin ring dynamics in the budding yeast.Microbiology (Reading). 2001 Jun;147(Pt 6):1437-1450. doi: 10.1099/00221287-147-6-1437. Microbiology (Reading). 2001. PMID: 11390675
-
A novel role for the alcohol sensitive ring/PHD finger protein Asr1p in regulating cell cycle mediated by septin-dependent assembly in yeast.Biochem Biophys Res Commun. 2015 Feb 27;458(1):208-13. doi: 10.1016/j.bbrc.2015.01.113. Epub 2015 Jan 31. Biochem Biophys Res Commun. 2015. PMID: 25646695
-
Architecture, remodeling, and functions of the septin cytoskeleton.Cytoskeleton (Hoboken). 2019 Jan;76(1):7-14. doi: 10.1002/cm.21475. Epub 2018 Aug 2. Cytoskeleton (Hoboken). 2019. PMID: 29979831 Free PMC article. Review.
-
The TORC2-Dependent Signaling Network in the Yeast Saccharomyces cerevisiae.Biomolecules. 2017 Sep 5;7(3):66. doi: 10.3390/biom7030066. Biomolecules. 2017. PMID: 28872598 Free PMC article. Review.
Cited by
-
Sequential recruitment of F-BAR proteins controls cytoskeletal crosstalk at the yeast bud neck.Curr Biol. 2025 Feb 3;35(3):574-590.e10. doi: 10.1016/j.cub.2024.12.011. Epub 2025 Jan 10. Curr Biol. 2025. PMID: 39798561
-
The final cut: cell polarity meets cytokinesis at the bud neck in S. cerevisiae.Cell Mol Life Sci. 2016 Aug;73(16):3115-36. doi: 10.1007/s00018-016-2220-3. Epub 2016 Apr 16. Cell Mol Life Sci. 2016. PMID: 27085703 Free PMC article. Review.
-
Septin-Associated Protein Kinases in the Yeast Saccharomyces cerevisiae.Front Cell Dev Biol. 2016 Nov 1;4:119. doi: 10.3389/fcell.2016.00119. eCollection 2016. Front Cell Dev Biol. 2016. PMID: 27847804 Free PMC article. Review.
-
Identification of Cell Cycle Dependent Interaction Partners of the Septins by Quantitative Mass Spectrometry.PLoS One. 2016 Feb 12;11(2):e0148340. doi: 10.1371/journal.pone.0148340. eCollection 2016. PLoS One. 2016. PMID: 26871441 Free PMC article.
-
Septin Organization and Functions in Budding Yeast.Front Cell Dev Biol. 2016 Nov 3;4:123. doi: 10.3389/fcell.2016.00123. eCollection 2016. Front Cell Dev Biol. 2016. PMID: 27857941 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
