

Neural circuit basis of visuo-spatial working memory precision: a computational and behavioral study
- PMID: 26180122
- PMCID: PMC4571767
- DOI: 10.1152/jn.00362.2015
Neural circuit basis of visuo-spatial working memory precision: a computational and behavioral study
Erratum in
-
CORRIGENDUM.J Neurophysiol. 2019 Oct 1;122(4):1843. doi: 10.1152/z9k-5270-corr.2019. J Neurophysiol. 2019. PMID: 31617797 Free PMC article. No abstract available.
Abstract
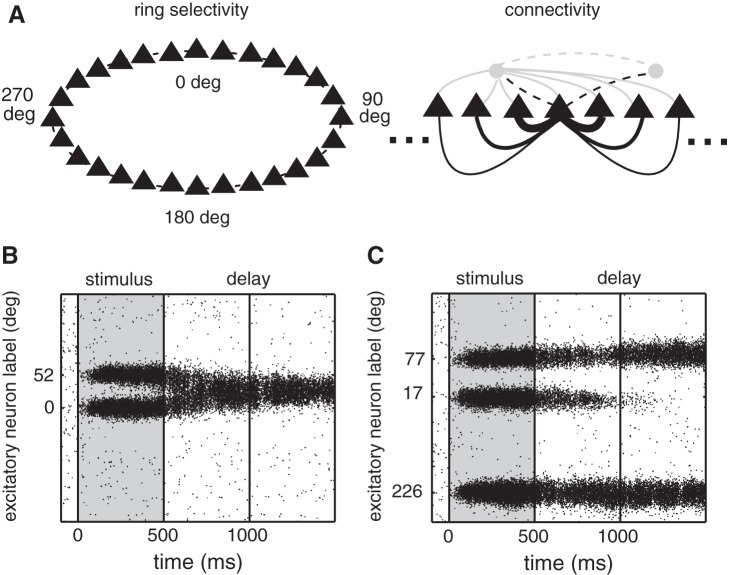
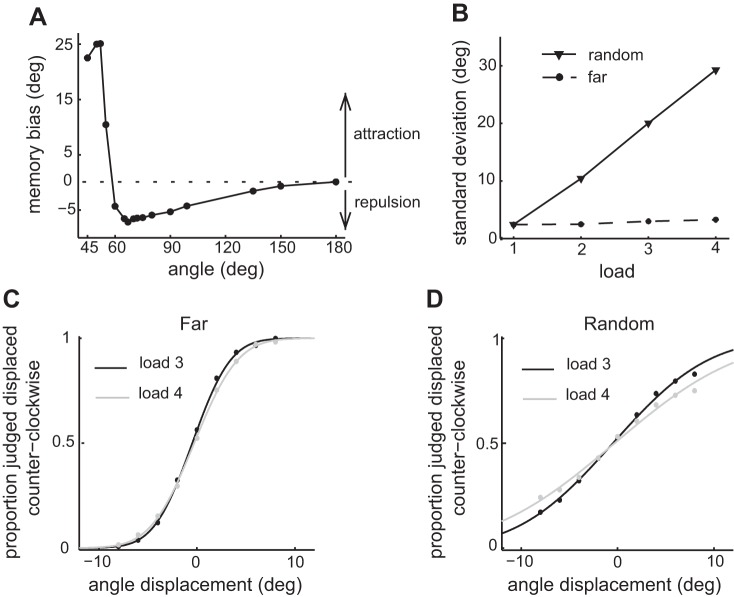
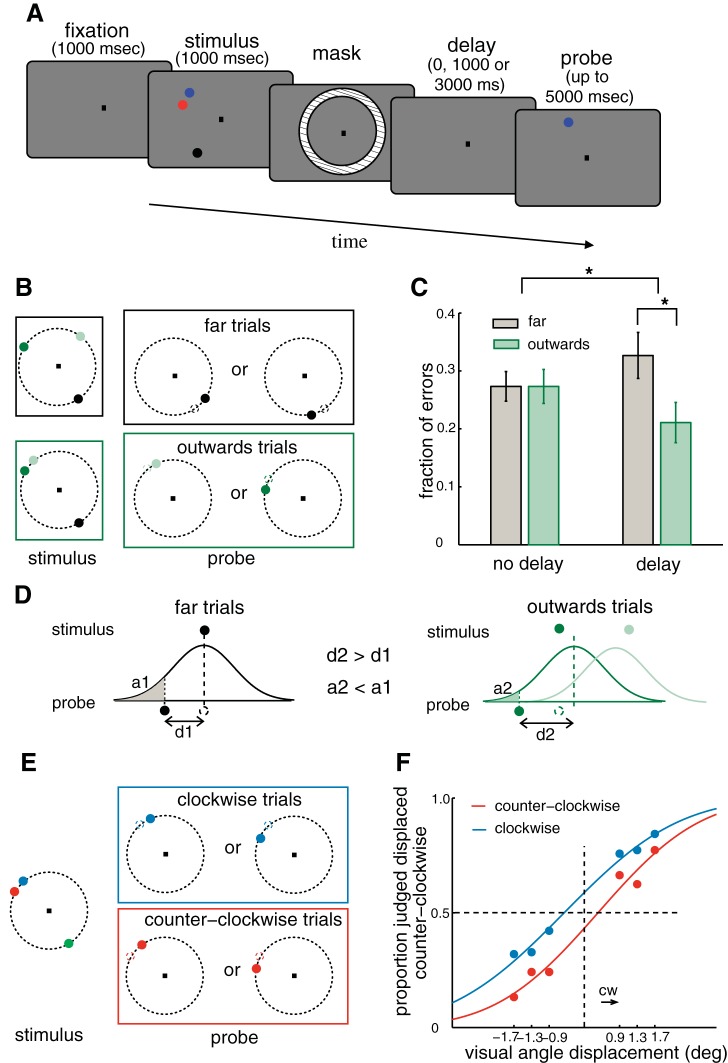
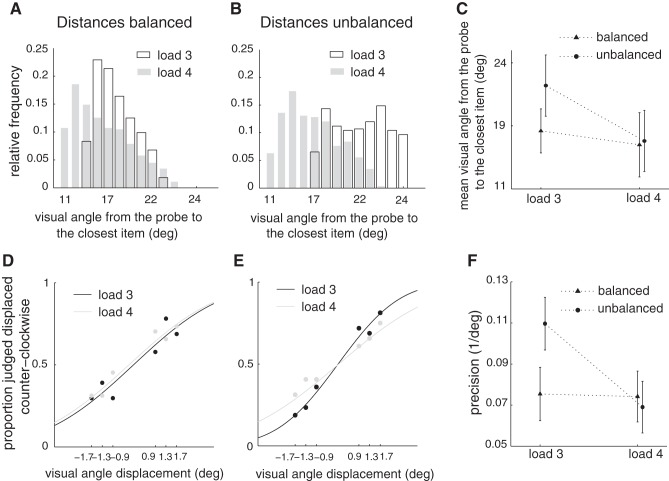
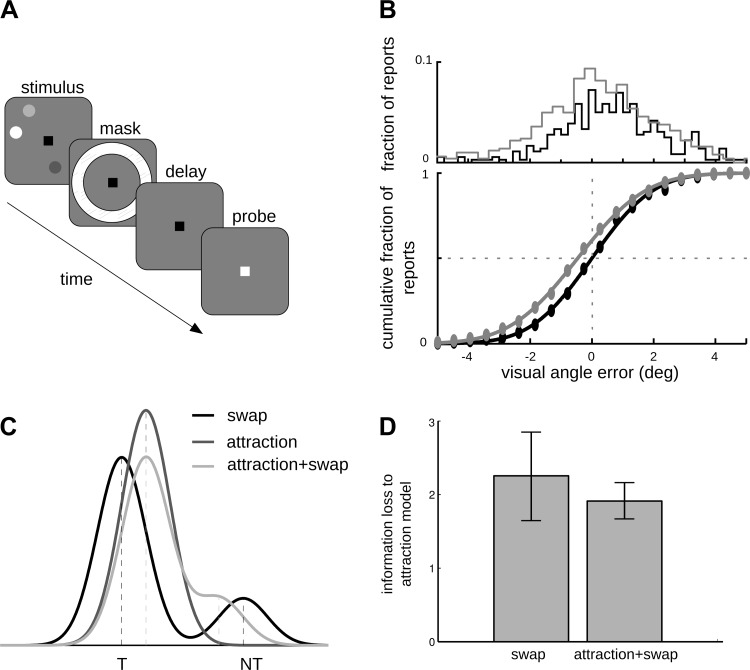
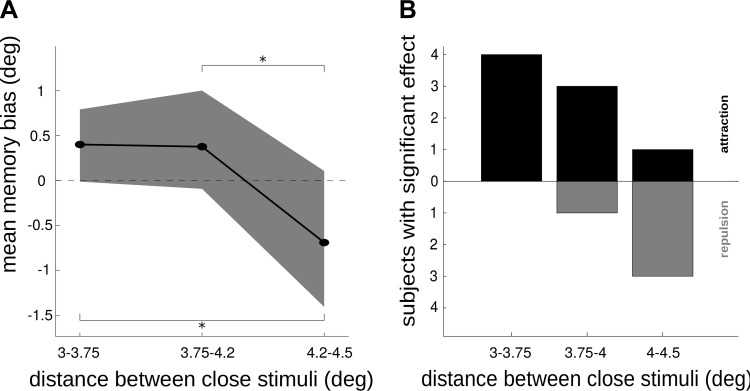
The amount of information that can be retained in working memory (WM) is limited. Limitations of WM capacity have been the subject of intense research, especially in trying to specify algorithmic models for WM. Comparatively, neural circuit perspectives have barely been used to test WM limitations in behavioral experiments. Here we used a neuronal microcircuit model for visuo-spatial WM (vsWM) to investigate memory of several items. The model assumes that there is a topographic organization of the circuit responsible for spatial memory retention. This assumption leads to specific predictions, which we tested in behavioral experiments. According to the model, nearby locations should be recalled with a bias, as if the two memory traces showed attraction or repulsion during the delay period depending on distance. Another prediction is that the previously reported loss of memory precision for an increasing number of memory items (memory load) should vanish when the distances between items are controlled for. Both predictions were confirmed experimentally. Taken together, our findings provide support for a topographic neural circuit organization of vsWM, they suggest that interference between similar memories underlies some WM limitations, and they put forward a circuit-based explanation that reconciles previous conflicting results on the dependence of WM precision with load.
Keywords: attractor model; capacity; precision; short-term memory; working memory.
Copyright © 2015 the American Physiological Society.
Figures
References
-
- Akaike H. A new look at the statistical model identification. IEEE Trans Automat Contr 19: 716–723, 1974.
-
- Alvarez G, Cavanagh P. The capacity of visual short-term memory is set both by visual information load and by number of objects. Psychol Sci 15: 106–111, 2004. - PubMed
-
- Amari SI. Dynamics of pattern formation in lateral-inhibition type neural fields. Biol Cybern 27: 77–87, 1977. - PubMed
-
- Baddeley A. Working Memory. New York: Oxford Univ. Press, 1986.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources