

Zebrafish Models for the Mechanosensory Hair Cell Dysfunction in Usher Syndrome 3 Reveal That Clarin-1 Is an Essential Hair Bundle Protein
- PMID: 26180195
- PMCID: PMC4502259
- DOI: 10.1523/JNEUROSCI.1096-15.2015
Zebrafish Models for the Mechanosensory Hair Cell Dysfunction in Usher Syndrome 3 Reveal That Clarin-1 Is an Essential Hair Bundle Protein
Abstract
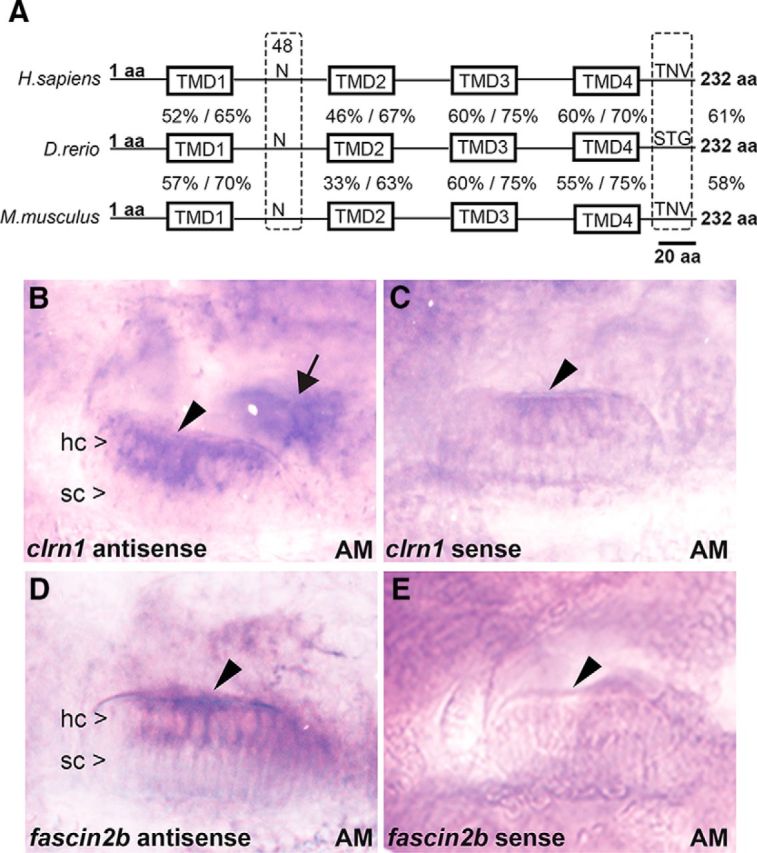
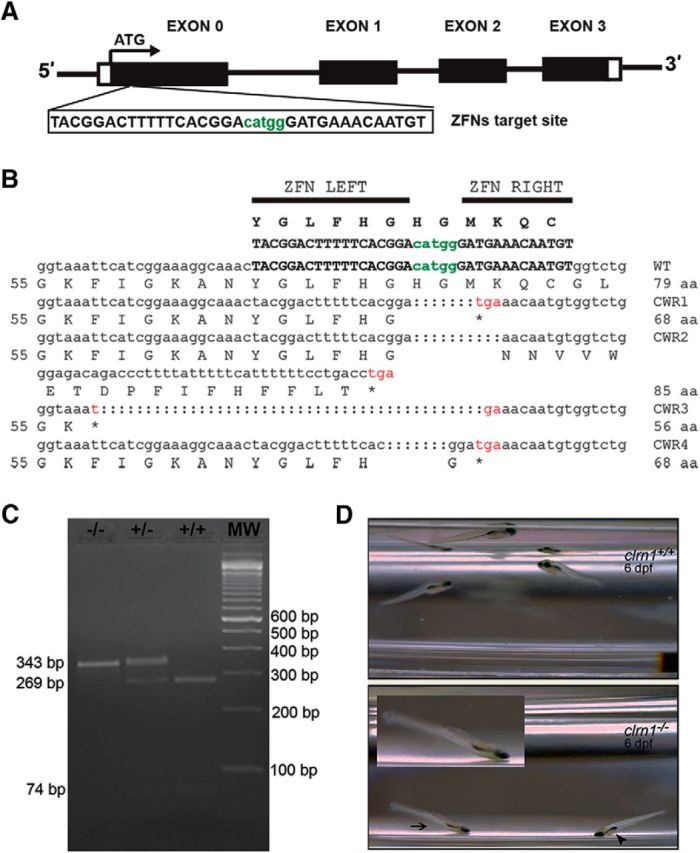
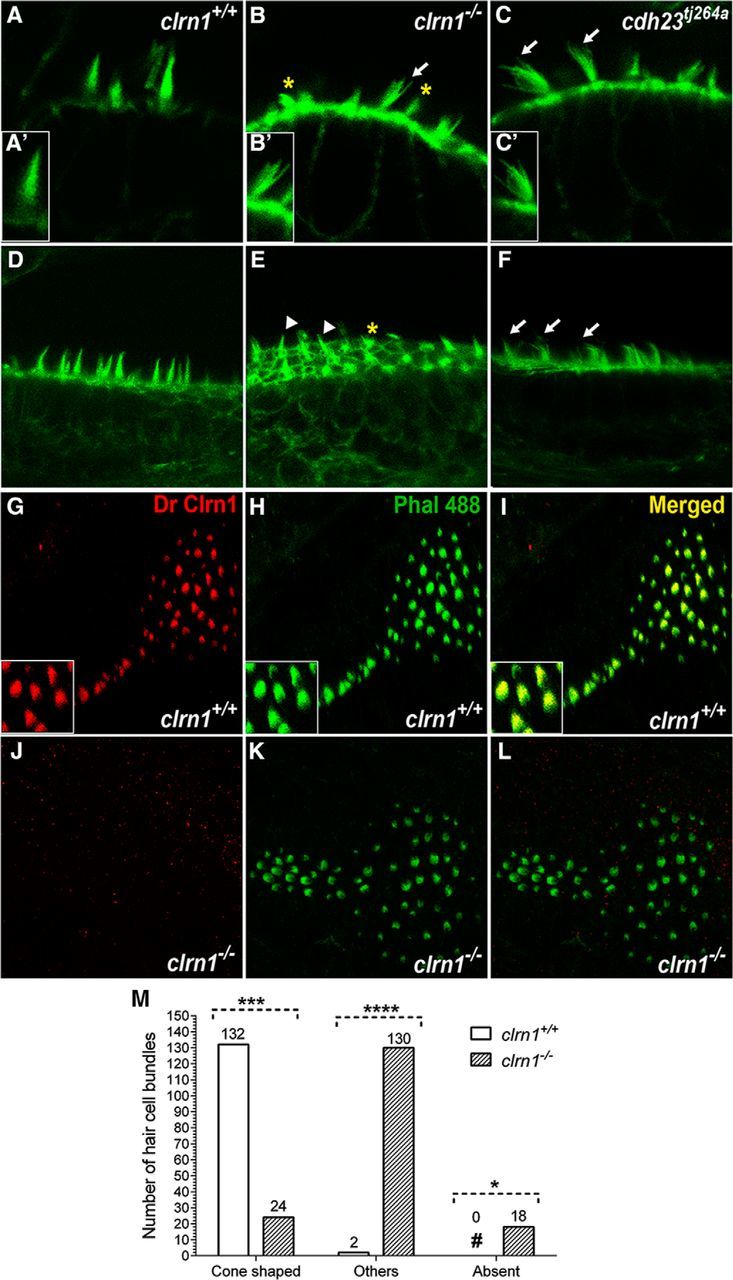
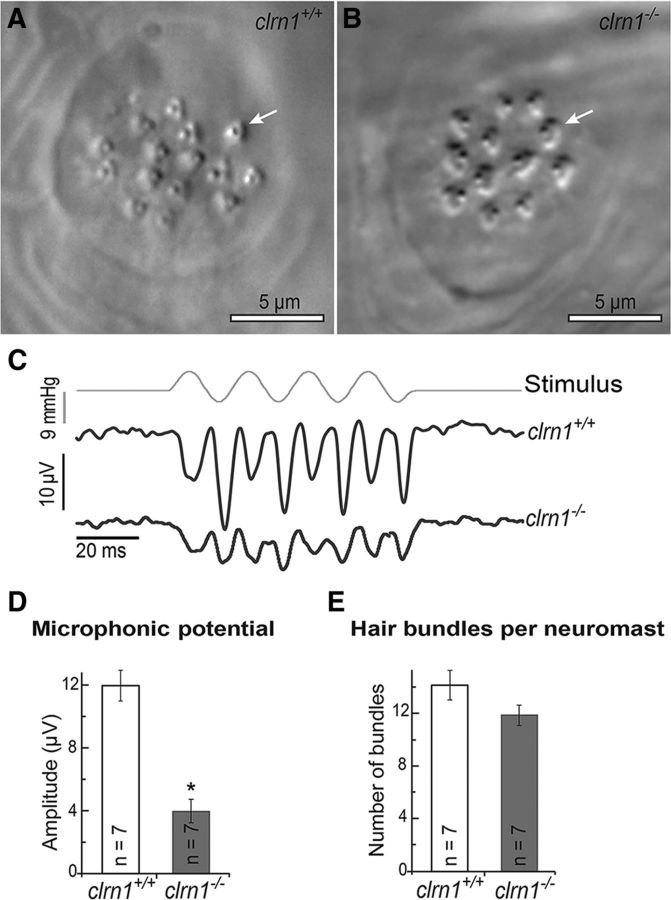
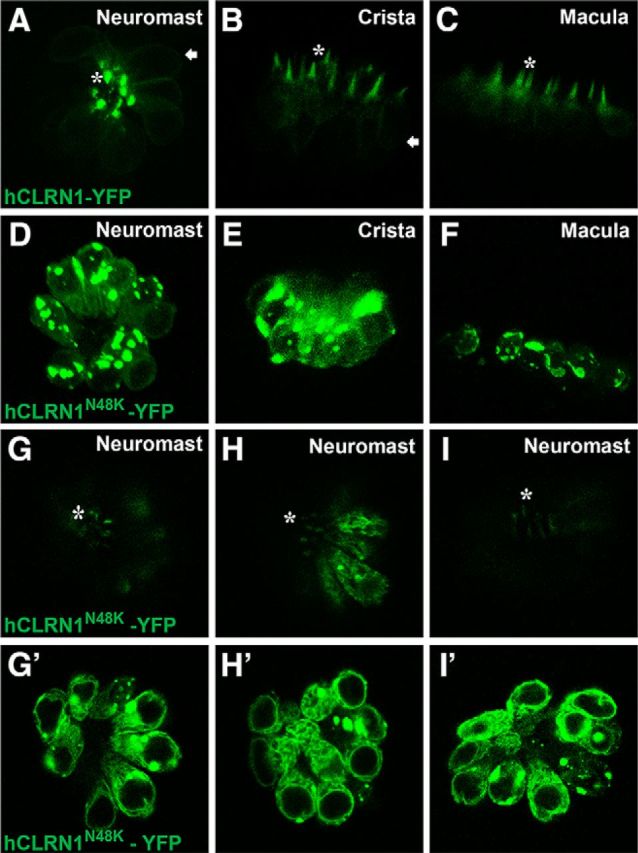
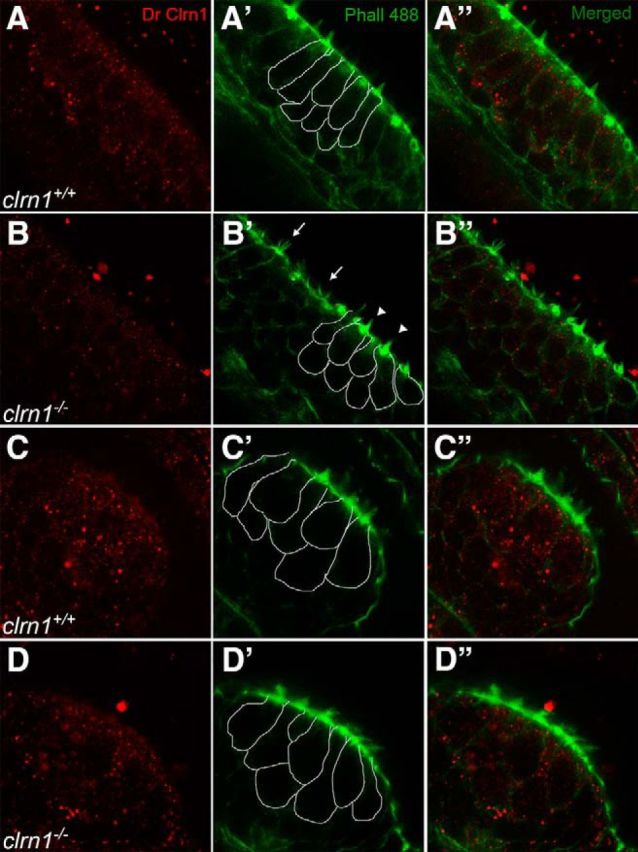
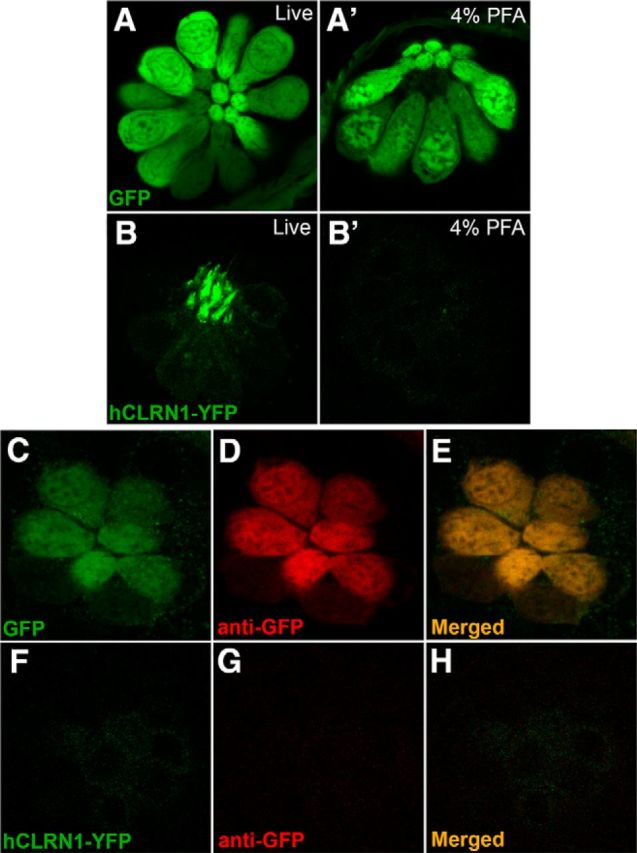
Usher syndrome type III (USH3) is characterized by progressive loss of hearing and vision, and varying degrees of vestibular dysfunction. It is caused by mutations that affect the human clarin-1 protein (hCLRN1), a member of the tetraspanin protein family. The missense mutation CLRN1(N48K), which affects a conserved N-glycosylation site in hCLRN1, is a common causative USH3 mutation among Ashkenazi Jews. The affected individuals hear at birth but lose that function over time. Here, we developed an animal model system using zebrafish transgenesis and gene targeting to provide an explanation for this phenotype. Immunolabeling demonstrated that Clrn1 localized to the hair cell bundles (hair bundles). The clrn1 mutants generated by zinc finger nucleases displayed aberrant hair bundle morphology with diminished function. Two transgenic zebrafish that express either hCLRN1 or hCLRN1(N48K) in hair cells were produced to examine the subcellular localization patterns of wild-type and mutant human proteins. hCLRN1 localized to the hair bundles similarly to zebrafish Clrn1; in contrast, hCLRN1(N48K) largely mislocalized to the cell body with a small amount reaching the hair bundle. We propose that this small amount of hCLRN1(N48K) in the hair bundle provides clarin-1-mediated function during the early stages of life; however, the presence of hCLRN1(N48K) in the hair bundle diminishes over time because of intracellular degradation of the mutant protein, leading to progressive loss of hair bundle integrity and hair cell function. These findings and genetic tools provide an understanding and path forward to identify therapies to mitigate hearing loss linked to the CLRN1 mutation.
Significance statement: Mutations in the clarin-1 gene affect eye and ear function in humans. Individuals with the CLRN1(N48K) mutation are born able to hear but lose that function over time. Here, we develop an animal model system using zebrafish transgenesis and gene targeting to provide an explanation for this phenotype. This approach illuminates the role of clarin-1 and the molecular mechanism linked to the CLRN1(N48K) mutation in sensory hair cells of the inner ear. Additionally, the investigation provided an in vivo model to guide future drug discovery to rescue the hCLRN1(N48K) in hair cells.
Keywords: clarin-1; hair cells; hearing.
Copyright © 2015 the authors 0270-6474/15/3510188-14$15.00/0.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Adato A, Vreugde S, Joensuu T, Avidan N, Hamalainen R, Belenkiy O, Olender T, Bonne-Tamir B, Ben-Asher E, Espinos C, Millán JM, Lehesjoki AE, Flannery JG, Avraham KB, Pietrokovski S, Sankila EM, Beckmann JS, Lancet D. USH3A transcripts encode clarin-1, a four-transmembrane-domain protein with a possible role in sensory synapses. Eur J Hum Genet. 2002;10:339–350. doi: 10.1038/sj.ejhg.5200831. - DOI - PubMed
-
- Adato A, Lefèvre G, Delprat B, Michel V, Michalski N, Chardenoux S, Weil D, El-Amraoui A, Petit C. Usherin, the defective protein in Usher syndrome type IIA, is likely to be a component of interstereocilia ankle links in the inner ear sensory cells. Hum Mol Genet. 2005;14:3921–3932. doi: 10.1093/hmg/ddi416. - DOI - PubMed
-
- Boëda B, El-Amraoui A, Bahloul A, Goodyear R, Daviet L, Blanchard S, Perfettini I, Fath KR, Shorte S, Reiners J, Houdusse A, Legrain P, Wolfrum U, Richardson G, Petit C. Myosin VIIa, harmonin and cadherin 23, three Usher I gene products that cooperate to shape the sensory hair cell bundle. EMBO J. 2002;21:6689–6699. doi: 10.1093/emboj/cdf689. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases