

Metabolism and Fitness of Urinary Tract Pathogens
- PMID: 26185076
- PMCID: PMC4510461
- DOI: 10.1128/microbiolspec.MBP-0016-2015
Metabolism and Fitness of Urinary Tract Pathogens
Abstract
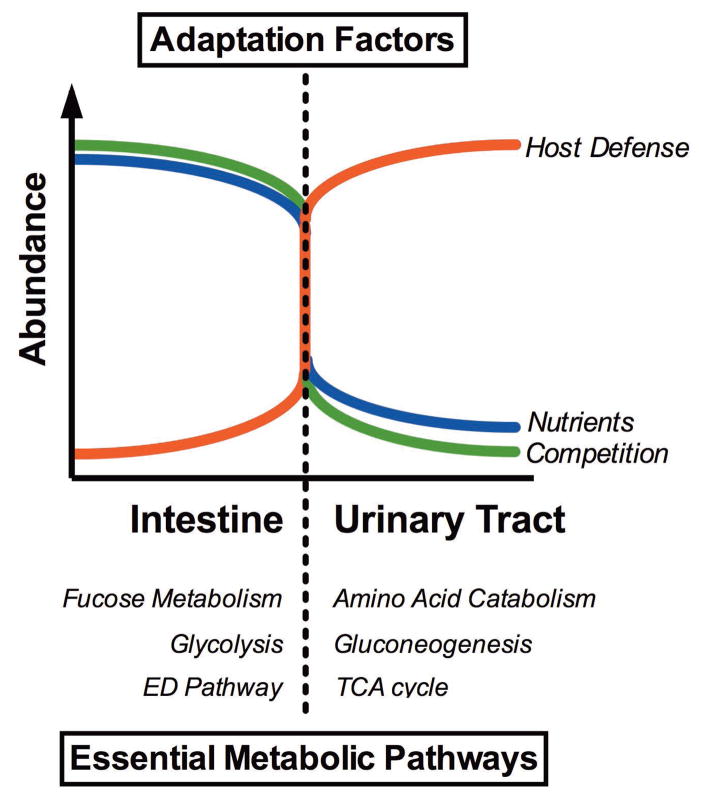
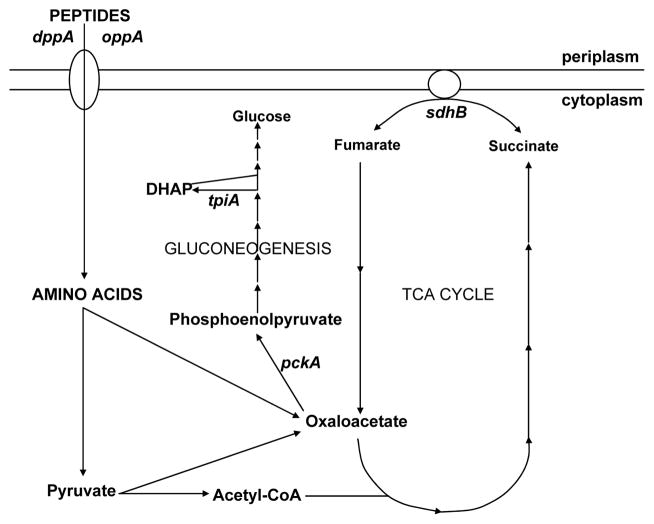
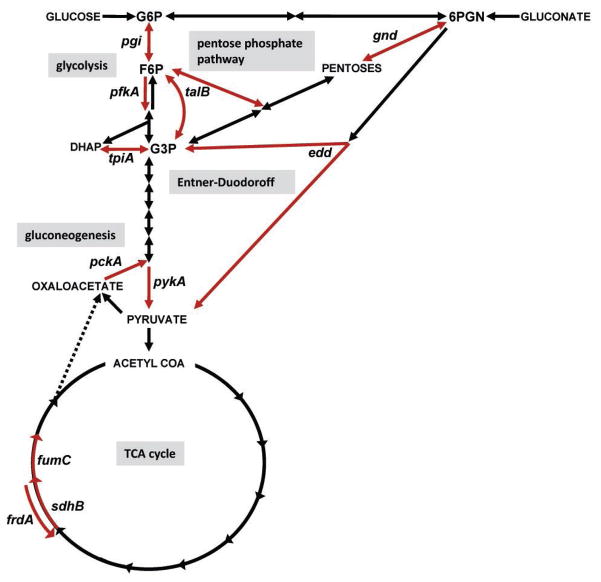
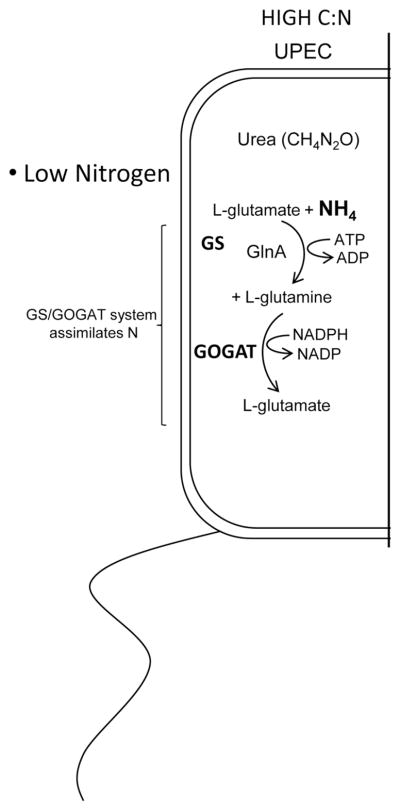
Among common infections, urinary tract infections (UTI) are the most frequently diagnosed urologic disease. The majority of UTIs are caused by uropathogenic Escherichia coli. The primary niche occupied by E. coli is the lower intestinal tract of mammals, where it resides as a beneficial component of the commensal microbiota. Although it is well-known that E. coli resides in the human intestine as a harmless commensal, specific strains or pathotypes have the potential to cause a wide spectrum of intestinal and diarrheal diseases. In contrast, extraintestinal E. coli pathotypes reside harmlessly in the human intestinal microenvironment but, upon access to sites outside of the intestine, become a major cause of human morbidity and mortality as a consequence of invasive UTI (pyelonephritis, bacteremia, or septicemia). Thus, extraintestinal pathotypes like uropathogenic E. coli (UPEC) possess an enhanced ability to cause infection outside of the intestinal tract and colonize the urinary tract, the bloodstream, or cerebrospinal fluid of human hosts. Due to the requirement for these E. coli to replicate in and colonize both the intestine and extraintestinal environments, we posit that physiology and metabolism of UPEC strains is paramount. Here we discuss that the ability to survive in the urinary tract depends as much on bacterial physiology and metabolism as it does on the well-considered virulence determinants.
Figures
References
-
- Foxman B, Barlow R, D’Arcy H, Gillespie B, Sobel JD. Urinary tract infection: self-reported incidence and associated costs. Ann Epidemiol. 2000;10:509–515. - PubMed
-
- Litwin MS, Saigal CS, Yano EM, Avila C, Geschwind SA, Hanley JM, Joyce GF, Madison R, Pace J, Polich SM, Wang M. Urologic diseases in America Project: analytical methods and principal findings. J Urol. 2005;173:933–937. - PubMed
-
- Kaper JB, Nataro JP, Mobley HL. Pathogenic Escherichia coli. Nat Rev Microbiol. 2004;2:123–140. - PubMed
-
- Touchon M, Hoede C, Tenaillon O, Barbe V, Baeriswyl S, Bidet P, Bingen E, Bonacorsi S, Bouchier C, Bouvet O, Calteau A, Chiapello H, Clermont O, Cruveiller S, Danchin A, Diard M, Dossat C, Karoui ME, Frapy E, Garry L, Ghigo JM, Gilles AM, Johnson J, Le Bouguenec C, Lescat M, Mangenot S, Martinez-Jehanne V, Matic I, Nassif X, Oztas S, Petit MA, Pichon C, Rouy Z, Ruf CS, Schneider D, Tourret J, Vacherie B, Vallenet D, Medigue C, Rocha EP, Denamur E. Organised genome dynamics in the Escherichia coli species results in highly diverse adaptive paths. PLoS Genet. 2009;5:e1000344. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
