

Impaired mitochondrial energy supply coupled to increased H2O2 emission under energy/redox stress leads to myocardial dysfunction during Type I diabetes
- PMID: 26186741
- PMCID: PMC6433379
- DOI: 10.1042/CS20150204
Impaired mitochondrial energy supply coupled to increased H2O2 emission under energy/redox stress leads to myocardial dysfunction during Type I diabetes
Abstract
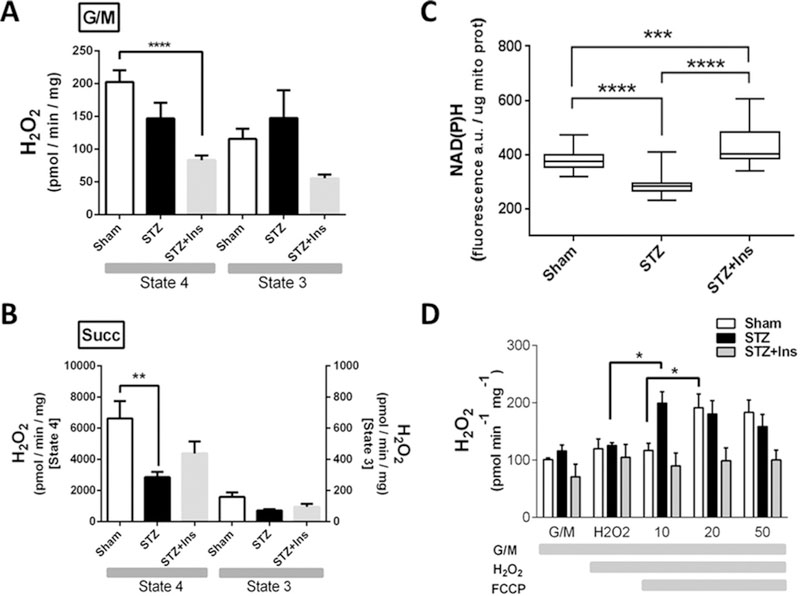
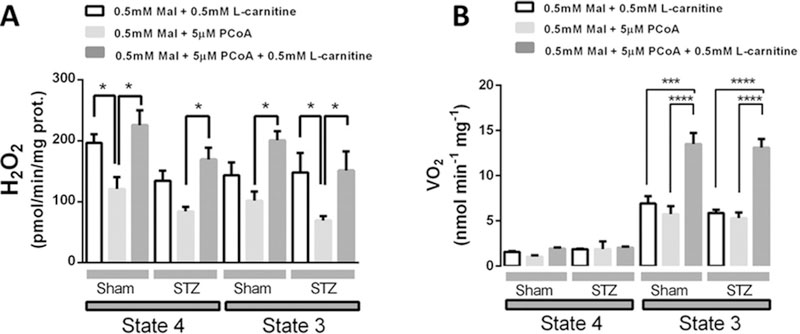
In Type I diabetic (T1DM) patients, both peaks of hyperglycaemia and increased sympathetic tone probably contribute to impair systolic and diastolic function. However, how these stressors eventually alter cardiac function during T1DM is not fully understood. In the present study, we hypothesized that impaired mitochondrial energy supply and excess reactive oxygen species (ROS) emission is centrally involved in T1DM cardiac dysfunction due to metabolic/redox stress and aimed to determine the mitochondrial sites implicated in these alterations. To this end, we used isolated myocytes and mitochondria from Sham and streptozotocin (STZ)-induced T1DM guinea pigs (GPs), untreated or treated with insulin. Relative to controls, T1DM myocytes exhibited higher oxidative stress when challenged with high glucose (HG) combined with β-adrenergic stimulation [via isoprenaline (isoproterenol) (ISO)], leading to contraction/relaxation deficits. T1DM mitochondria had decreased respiration with complex II and IV substrates and markedly lower ADP phosphorylation rates and higher H2O2 emission when challenged with oxidants to mimic the more oxidized redox milieu present in HG + ISO-treated cardiomyocytes. Since in T1DM hearts insulin-sensitivity is preserved and a glucose-to-fatty acid (FA) shift occurs, we next tested whether insulin therapy or acute palmitate (Palm) infusion prevents HG + ISO-induced cardiac dysfunction. We found that insulin rescued proper cardiac redox balance, but not mitochondrial respiration or contractile performance. Conversely, Palm restored redox balance and preserved myocyte function. Thus, stressors such as peaks of HG and adrenergic hyperactivity impair mitochondrial respiration, hampering energy supply while exacerbating ROS emission. Our study suggests that an ideal therapeutic measure to treat metabolically/redox-challenged T1DM hearts should concomitantly correct energetic and redox abnormalities to fully maintain cardiac function.
Keywords: adrenergic stimulation; calcium transient; contractility; diabetes; energetic transitions; glutathione; hyperglycaemia; redox balance.
© 2015 Authors; published by Portland Press Limited.
Figures




References
-
- Jyotsna VP, Sahoo A, Sreenivas V and Deepak KK (2009) Prevalence and pattern of cardiac autonomic dysfunction in newly detected type 2 diabetes mellitus. Diabetes Res. Clin. Pract. 83, 83–88 - PubMed
-
- Choi SW, Benzie IF, Ma SW, Strain JJ and Hannigan BM (2008) Acute hyperglycemia and oxidative stress: direct cause and effect? Free Radic. Biol. Med. 44, 1217–1231 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical