

Commentary: IL-4 and IL-13 receptors and signaling
- PMID: 26187331
- PMCID: PMC4546937
- DOI: 10.1016/j.cyto.2015.05.023
Commentary: IL-4 and IL-13 receptors and signaling
Abstract
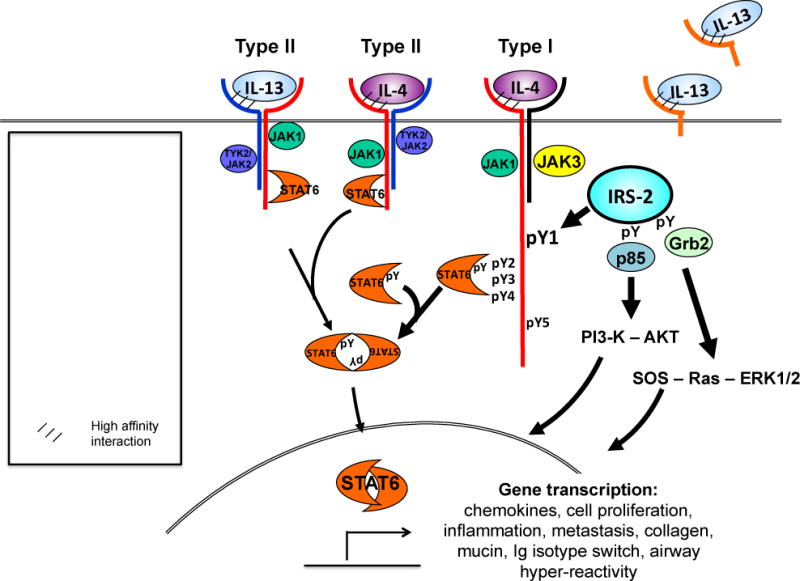
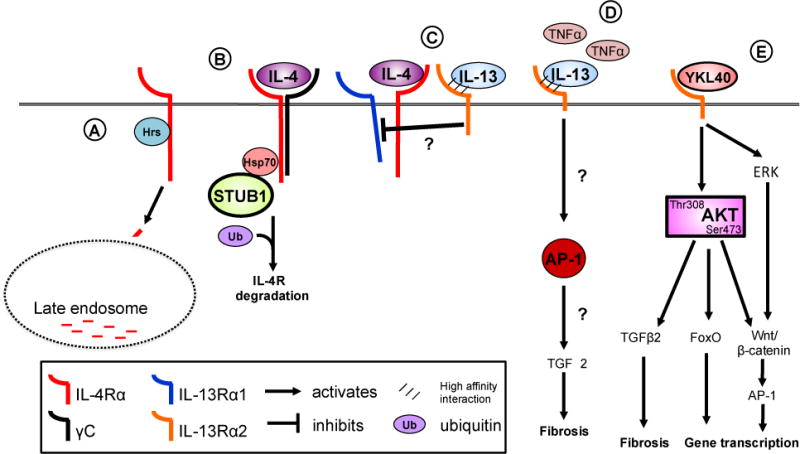
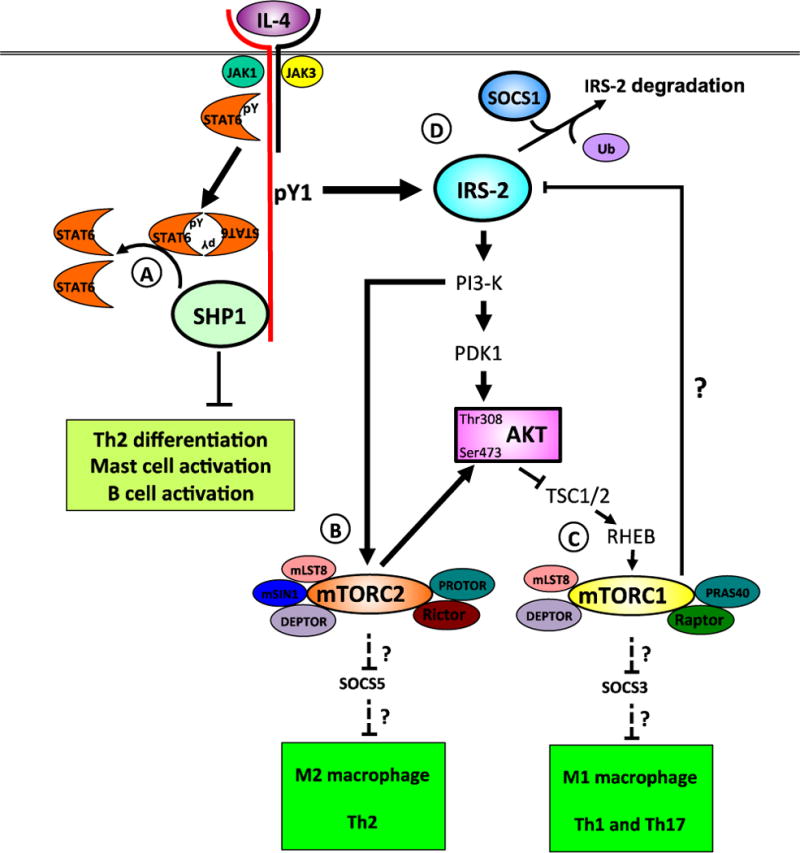
Interleukin (IL)-4 and IL-13 were discovered approximately 30years ago and were immediately linked to allergy and atopic diseases. Since then, new roles for IL-4 and IL-13 and their receptors in normal gestation, fetal development and neurological function and in the pathogenesis of cancer and fibrosis have been appreciated. Studying IL-4/-13 and their receptors has revealed important clues about cytokine biology and led to the development of numerous experimental therapeutics. Here we aim to highlight new discoveries and consolidate concepts in the field of IL-4 and IL-13 structure, receptor regulation, signaling and experimental therapeutics.
Keywords: Interleukin-13; Interleukin-4; Receptor; Regulation; Singling.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
References
-
- McKenzie Andrew NJ, Spits H, Eberl G. Innate Lymphoid Cells in Inflammation and Immunity. Immunity. 2014;41:366–374. - PubMed
-
- Hams E, Fallon PG. Innate type 2 cells and asthma. Current Opinion in Pharmacology. 2012;12:503–509. - PubMed
-
- Junttila IS, Creusot RJ, Moraga I, Bates DL, Wong MT, Alonso MN, Suhoski MM, Lupardus P, Meier-Schellersheim M, Engleman EG, Utz PJ, Fathman CG, Paul WE, Garcia KC. Redirecting cell-type specific cytokine responses with engineered interleukin-4 superkines. Nat Chem Biol. 2012;8:990–998. - PMC - PubMed
-
- Heller NM, Dasgupta P, Dorsey NJ, Chapoval SP, Keegan AD. The Type I and Type II Receptor Complexes for IL-4 and IL-13 Differentially Regulate Allergic Lung Inflammation, Allergic Diseases. Pereira C, editor. Highlights in the Clinic, Mechanisms and Treatment. 2012 http://www.intechopen.com/books/allergic-diseases-highlights-in-the-clin....
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
