

Mosquitoes Use Vision to Associate Odor Plumes with Thermal Targets
- PMID: 26190071
- PMCID: PMC4546539
- DOI: 10.1016/j.cub.2015.06.046
Mosquitoes Use Vision to Associate Odor Plumes with Thermal Targets
Abstract
All moving animals, including flies, sharks, and humans, experience a dynamic sensory landscape that is a function of both their trajectory through space and the distribution of stimuli in the environment. This is particularly apparent for mosquitoes, which use a combination of olfactory, visual, and thermal cues to locate hosts. Mosquitoes are thought to detect suitable hosts by the presence of a sparse CO₂ plume, which they track by surging upwind and casting crosswind. Upon approach, local cues such as heat and skin volatiles help them identify a landing site. Recent evidence suggests that thermal attraction is gated by the presence of CO₂, although this conclusion was based experiments in which the actual flight trajectories of the animals were unknown and visual cues were not studied. Using a three-dimensional tracking system, we show that rather than gating heat sensing, the detection of CO₂ actually activates a strong attraction to visual features. This visual reflex guides the mosquitoes to potential hosts where they are close enough to detect thermal cues. By experimentally decoupling the olfactory, visual, and thermal cues, we show that the motor reactions to these stimuli are independently controlled. Given that humans become visible to mosquitoes at a distance of 5-15 m, visual cues play a critical intermediate role in host localization by coupling long-range plume tracking to behaviors that require short-range cues. Rather than direct neural coupling, the separate sensory-motor reflexes are linked as a result of the interaction between the animal's reactions and the spatial structure of the stimuli in the environment.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
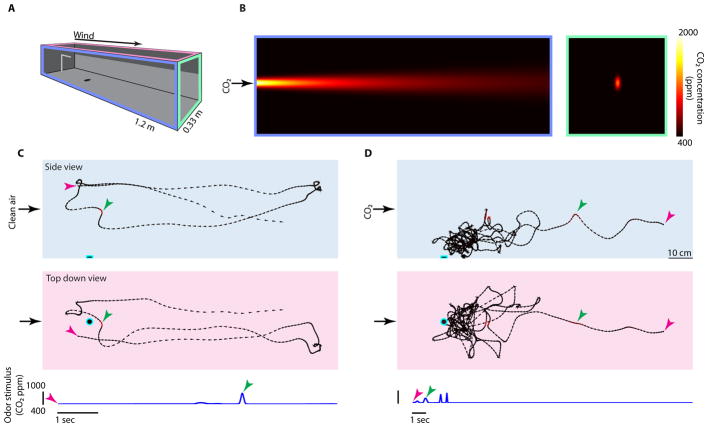
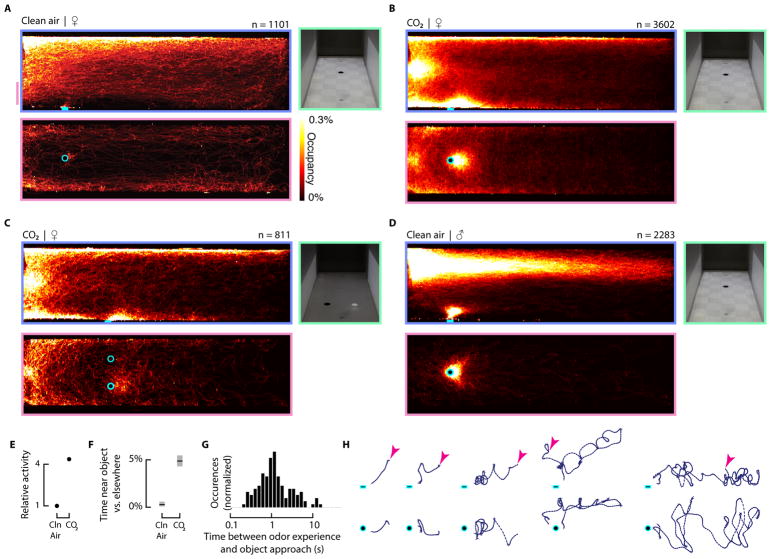
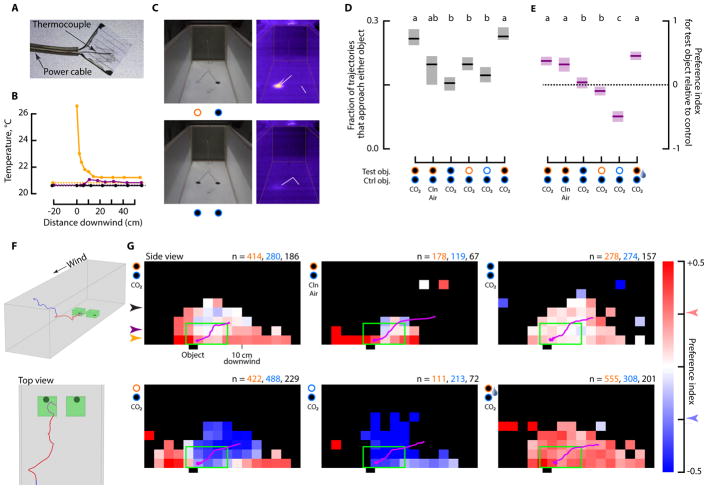
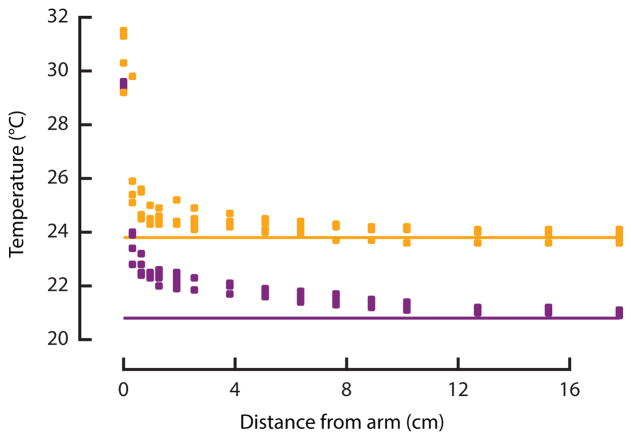
Comment in
-
Multi-Cue Integration: How Female Mosquitoes Locate a Human Host.Curr Biol. 2015 Sep 21;25(18):R793-5. doi: 10.1016/j.cub.2015.07.057. Curr Biol. 2015. PMID: 26394099
References
-
- Van Breugel F, Dickinson MH. Plume-tracking behavior of flying Drosophila emerges from a set of distinct sensory-motor reflexes. Curr Biol. 2014;24:274–86. - PubMed
-
- Budick SA, Dickinson MH. Free-flight responses of Drosophila melanogaster to attractive odors. J Exp Biol. 2006;209:3001–17. - PubMed
-
- Barrows WM. The reactions of the Pomace fly, Drosophila ampelophila loew, to odorous substances. J Exp Zool. 1907;4:515–537.
-
- Johnsen PB, Teeter JH. Behavioral responses of bonnethead sharks (Sphyrna tiburo) to controlled olfactory stimulation. Mar Freshw Behav Phy. 1985;11:283–291.
-
- Porter J, Craven B, Khan RM, Chang SJ, Kang I, Judkewitz B, Judkewicz B, Volpe J, Settles G, Sobel N. Mechanisms of scent-tracking in humans. Nat Neurosci. 2007;10:27–9. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
