

Stem cells are units of natural selection for tissue formation, for germline development, and in cancer development
- PMID: 26195745
- PMCID: PMC4517284
- DOI: 10.1073/pnas.1505464112
Stem cells are units of natural selection for tissue formation, for germline development, and in cancer development
Abstract
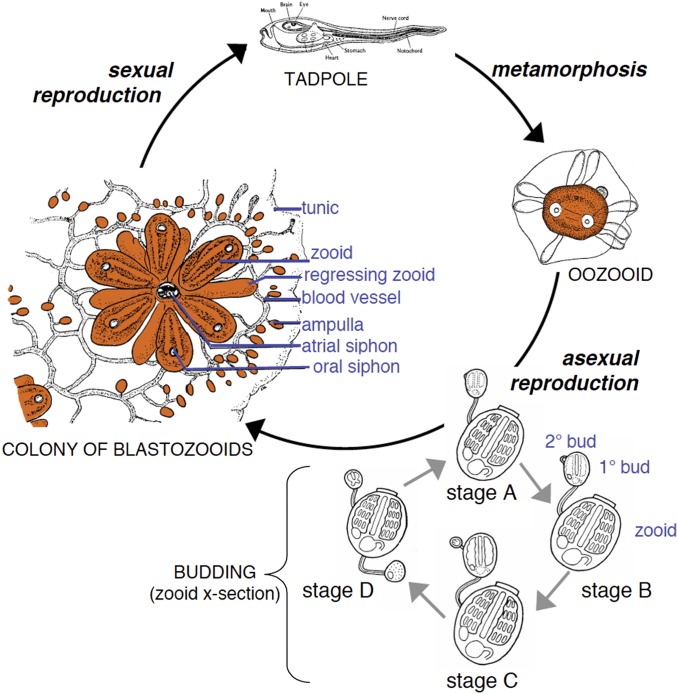
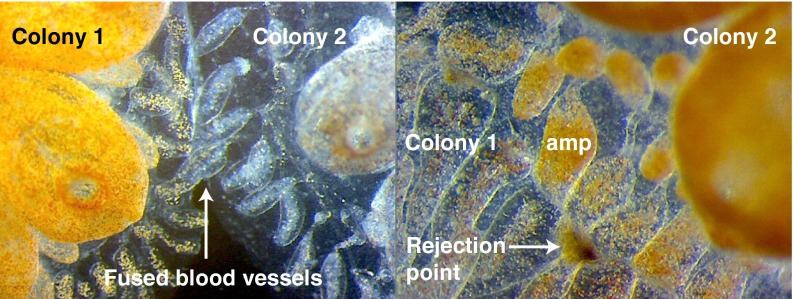
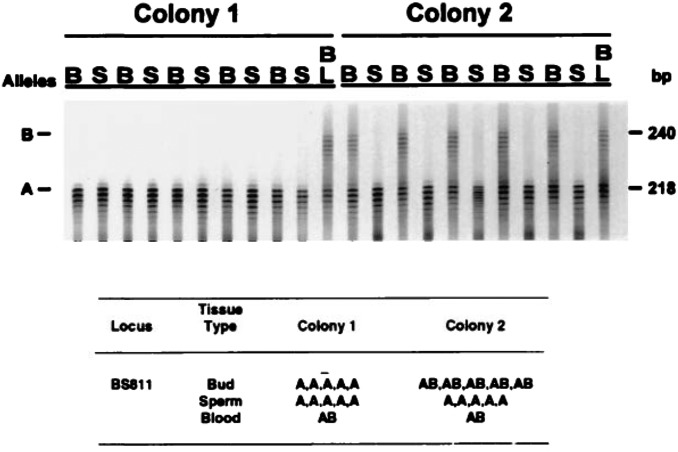
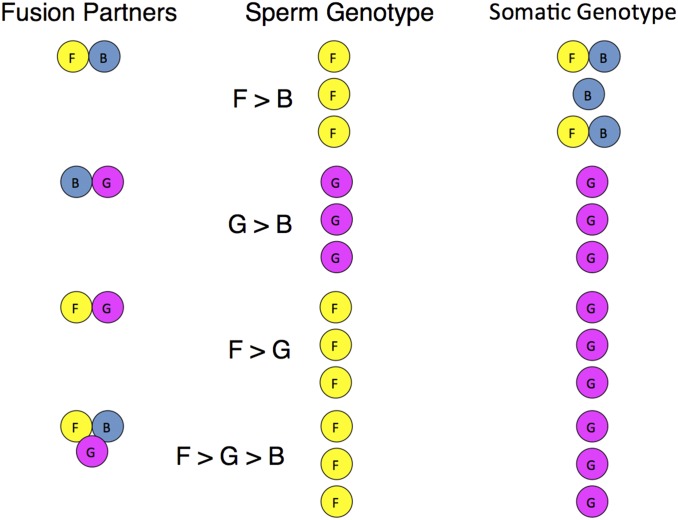
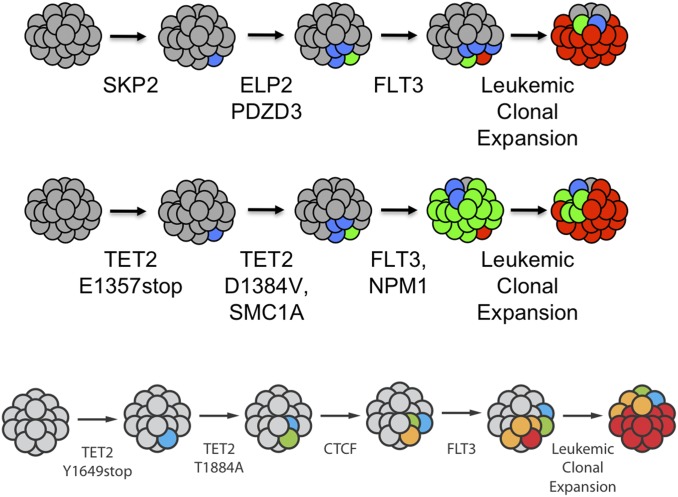
It is obvious that natural selection operates at the level of individuals and collections of individuals. Nearly two decades ago we showed that in multi-individual colonies of protochordate colonial tunicates sharing a blood circulation, there exists an exchange of somatic stem cells and germline stem cells, resulting in somatic chimeras and stem cell competitions for gonadal niches. Stem cells are unlike other cells in the body in that they alone self-renew, so that they form clones that are perpetuated for the life of the organism. Stem cell competitions have allowed the emergence of competitive somatic and germline stem cell clones. Highly successful germline stem cells usually outcompete less successful competitors both in the gonads of the genotype partner from which they arise and in the gonads of the natural parabiotic partners. Therefore, natural selection also operates at the level of germline stem cell clones. In the colonial tunicate Botryllus schlosseri the formation of natural parabionts is prevented by a single-locus highly polymorphic histocompatibility gene called Botryllus histocompatibility factor. This limits germline stem cell predation to kin, as the locus has hundreds of alleles. We show that in mice germline stem cells compete for gonad niches, and in mice and humans, blood-forming stem cells also compete for bone marrow niches. We show that the clonal progression from blood-forming stem cells to acute leukemias by successive genetic and epigenetic events in blood stem cells also involves competition and selection between clones and propose that this is a general theme in cancer.
Keywords: Botryllus schlosseri; cancer; evolution; natural selection; stem cell competition.
Conflict of interest statement
The author declares no conflict of interest.
Figures
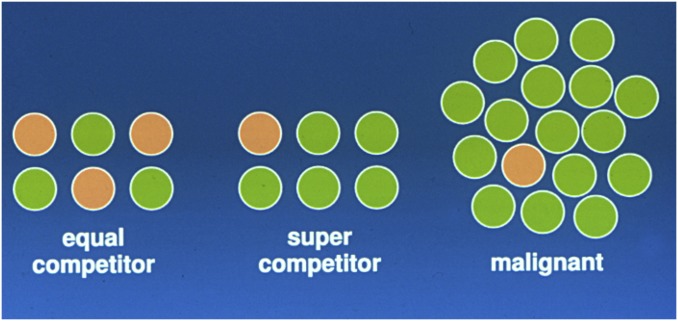
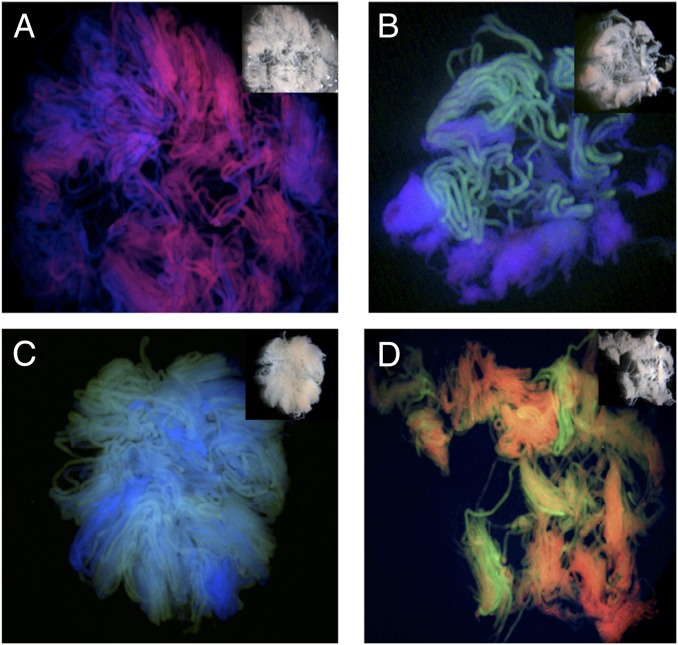
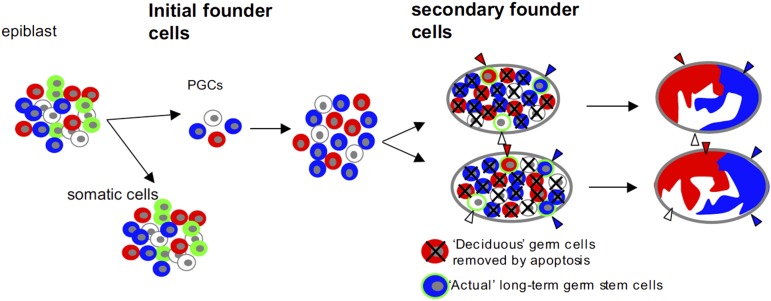
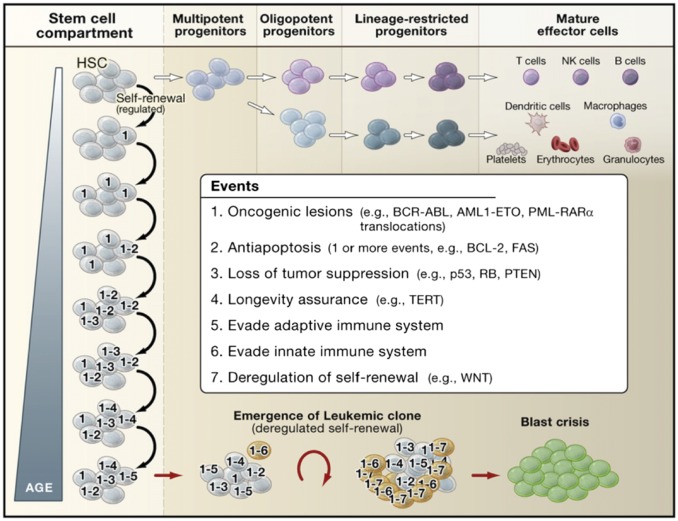
References
-
- Weissman IL. Stem cells: Units of development, units of regeneration, and units in evolution. Cell. 2000;100(1):157–168. - PubMed
-
- Laird DJ, Weissman IL. Telomerase maintained in self-renewing tissues during serial regeneration of the urochordate Botryllus schlosseri. Dev Biol. 2004;273(2):185–194. - PubMed
-
- Scofield VL, Schlumpberger JM, West LA, Weissman IL. Protochordate allorecognition is controlled by a MHC-like gene system. Nature. 1982;295(5849):499–502. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
