

Large-Scale Phenomics Identifies Primary and Fine-Tuning Roles for CRKs in Responses Related to Oxidative Stress
- PMID: 26197346
- PMCID: PMC4511522
- DOI: 10.1371/journal.pgen.1005373
Large-Scale Phenomics Identifies Primary and Fine-Tuning Roles for CRKs in Responses Related to Oxidative Stress
Abstract
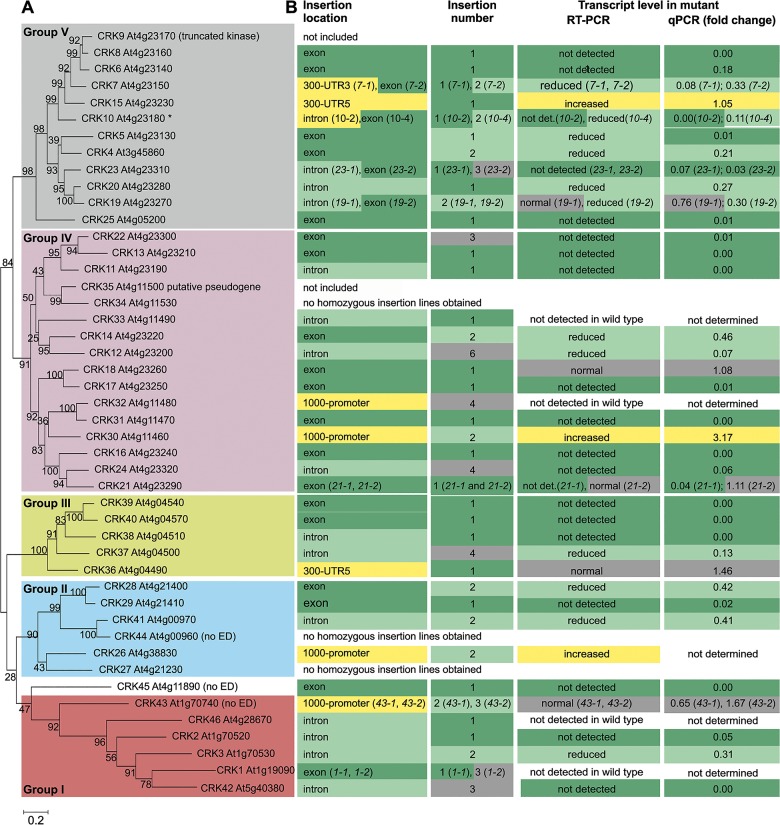
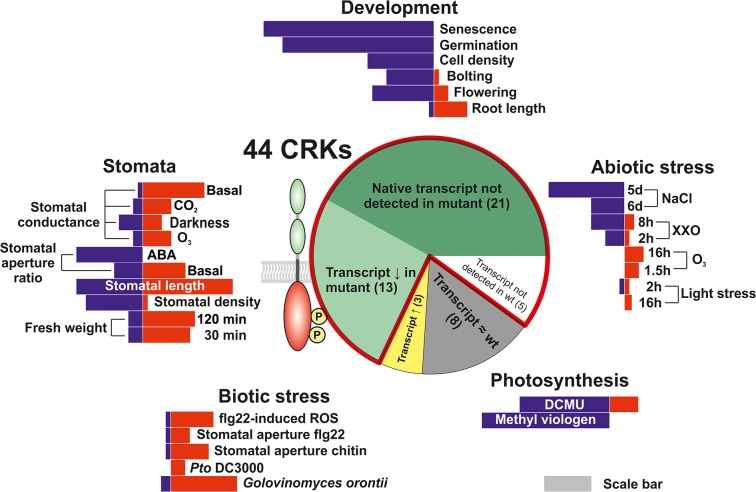
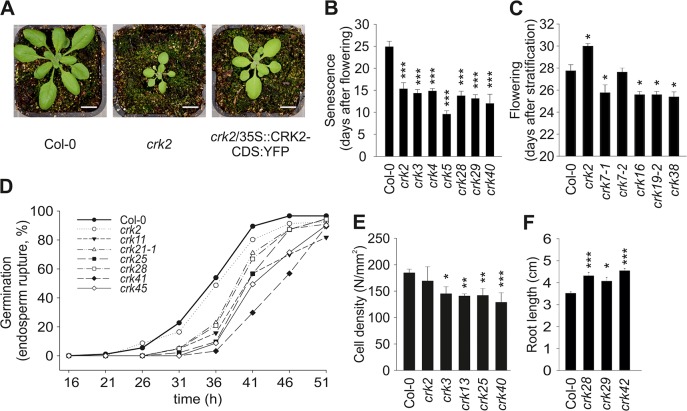
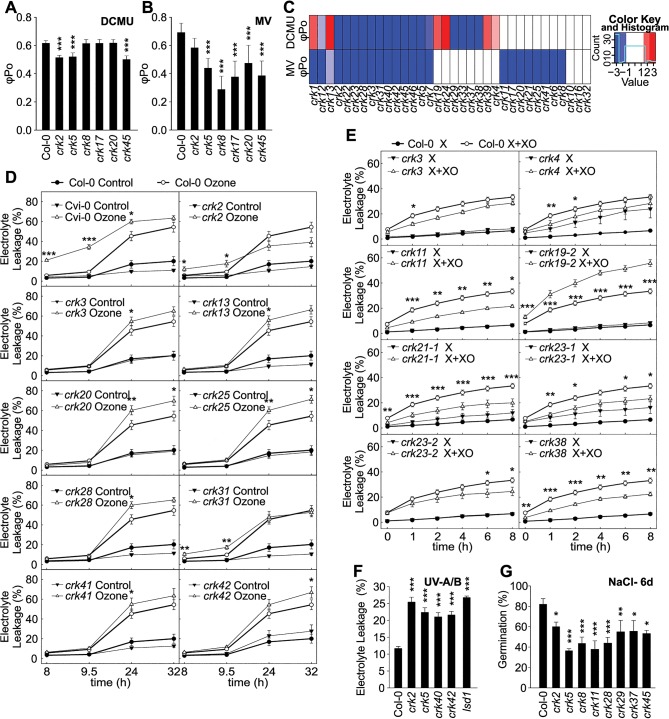
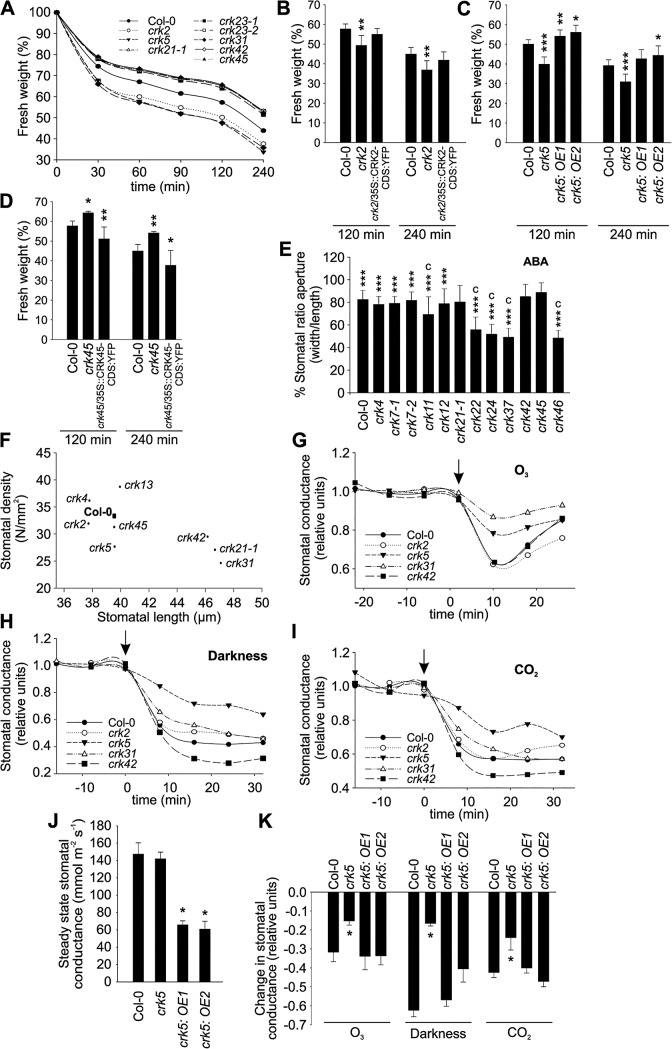
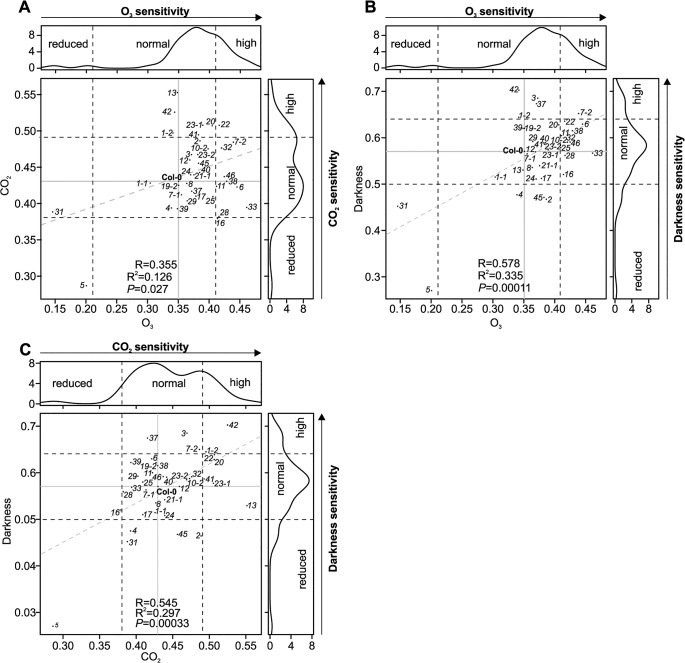
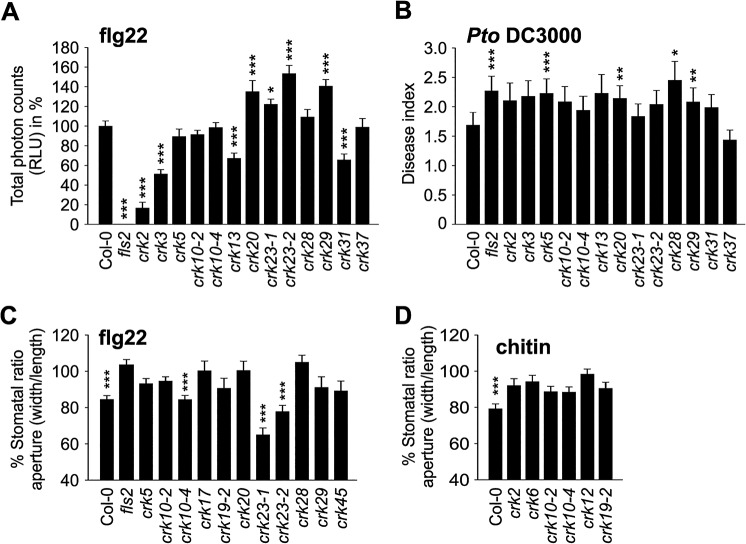
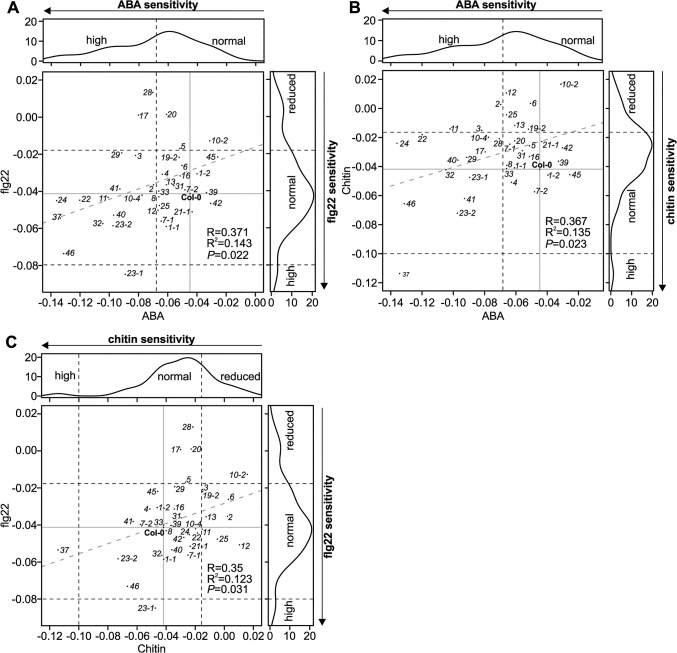
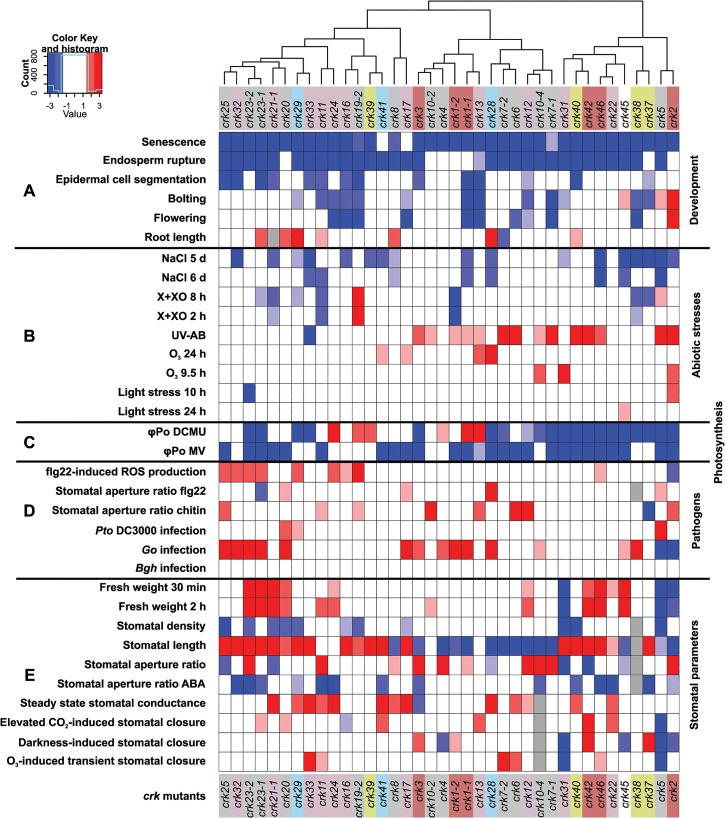
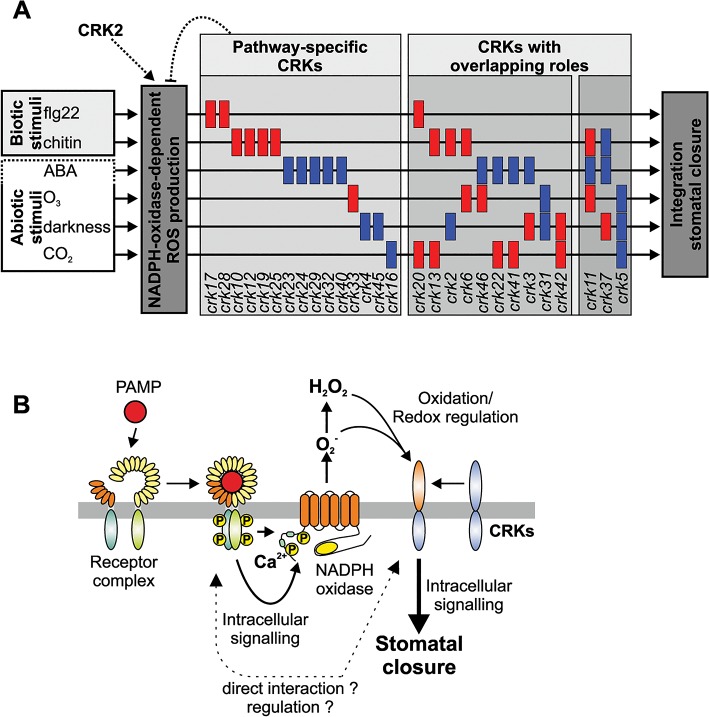
Cysteine-rich receptor-like kinases (CRKs) are transmembrane proteins characterized by the presence of two domains of unknown function 26 (DUF26) in their ectodomain. The CRKs form one of the largest groups of receptor-like protein kinases in plants, but their biological functions have so far remained largely uncharacterized. We conducted a large-scale phenotyping approach of a nearly complete crk T-DNA insertion line collection showing that CRKs control important aspects of plant development and stress adaptation in response to biotic and abiotic stimuli in a non-redundant fashion. In particular, the analysis of reactive oxygen species (ROS)-related stress responses, such as regulation of the stomatal aperture, suggests that CRKs participate in ROS/redox signalling and sensing. CRKs play general and fine-tuning roles in the regulation of stomatal closure induced by microbial and abiotic cues. Despite their great number and high similarity, large-scale phenotyping identified specific functions in diverse processes for many CRKs and indicated that CRK2 and CRK5 play predominant roles in growth regulation and stress adaptation, respectively. As a whole, the CRKs contribute to specificity in ROS signalling. Individual CRKs control distinct responses in an antagonistic fashion suggesting future potential for using CRKs in genetic approaches to improve plant performance and stress tolerance.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
Cysteine-rich receptor-like kinase CRK5 as a regulator of growth, development, and ultraviolet radiation responses in Arabidopsis thaliana.J Exp Bot. 2015 Jun;66(11):3325-37. doi: 10.1093/jxb/erv143. Epub 2015 May 11. J Exp Bot. 2015. PMID: 25969551 Free PMC article.
-
Transcriptional regulation of the CRK/DUF26 group of receptor-like protein kinases by ozone and plant hormones in Arabidopsis.BMC Plant Biol. 2010 May 25;10:95. doi: 10.1186/1471-2229-10-95. BMC Plant Biol. 2010. PMID: 20500828 Free PMC article.
-
Activation of hypersensitive cell death by pathogen-induced receptor-like protein kinases from Arabidopsis.Plant Mol Biol. 2004 Sep;56(2):271-83. doi: 10.1007/s11103-004-3381-2. Plant Mol Biol. 2004. PMID: 15604743
-
More questions than answers: insights into potential cysteine-rich receptor-like kinases redox signalling in Arabidopsis.Plant J. 2025 Apr;122(2):e70176. doi: 10.1111/tpj.70176. Plant J. 2025. PMID: 40300149 Free PMC article. Review.
-
Cysteine-rich receptor-like protein kinases: emerging regulators of plant stress responses.Trends Plant Sci. 2023 Jul;28(7):776-794. doi: 10.1016/j.tplants.2023.03.028. Epub 2023 Apr 26. Trends Plant Sci. 2023. PMID: 37105805 Review.
Cited by
-
A point mutation in the kinase domain of CRK10 leads to xylem vessel collapse and activation of defence responses in Arabidopsis.J Exp Bot. 2023 May 19;74(10):3104-3121. doi: 10.1093/jxb/erad080. J Exp Bot. 2023. PMID: 36869735 Free PMC article.
-
RNA N6-Methyladenosine Affects Copper-Induced Oxidative Stress Response in Arabidopsis thaliana.Noncoding RNA. 2024 Jan 19;10(1):8. doi: 10.3390/ncrna10010008. Noncoding RNA. 2024. PMID: 38392963 Free PMC article.
-
ROS Regulation of Polar Growth in Plant Cells.Plant Physiol. 2016 Jul;171(3):1593-605. doi: 10.1104/pp.16.00191. Epub 2016 May 4. Plant Physiol. 2016. PMID: 27208283 Free PMC article. Review.
-
CRK2 Enhances Salt Tolerance by Regulating Callose Deposition in Connection with PLDα1.Plant Physiol. 2019 Aug;180(4):2004-2021. doi: 10.1104/pp.19.00560. Epub 2019 May 22. Plant Physiol. 2019. PMID: 31118265 Free PMC article.
-
Explaining pre-emptive acclimation by linking information to plant phenotype.J Exp Bot. 2022 Sep 3;73(15):5213-5234. doi: 10.1093/jxb/erab537. J Exp Bot. 2022. PMID: 34915559 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
