

Review
doi: 10.1007/s12015-015-9597-5.
X Inactivation Lessons from Differentiating Mouse Embryonic Stem Cells
Affiliations
- PMID: 26198263
- PMCID: PMC4561061
- DOI: 10.1007/s12015-015-9597-5
Item in Clipboard
Review
X Inactivation Lessons from Differentiating Mouse Embryonic Stem Cells
Stem Cell Rev Rep.
2015 Oct.
Abstract
X chromosome inactivation (XCI) is the dosage compensation mechanism that evolved in female mammals to correct the genetic imbalance of X-linked genes between sexes. X chromosome inactivation occurs in early development when one of the two X chromosomes of females is nearly-completely silenced. Differentiating Embryonic Stem cells (ESC) are regarded as a useful tool to study XCI, since they recapitulate many events occurring during early development. In this review we aim to summarise the advances in the field and to discuss the close connection between cell differentiation and X chromosome inactivation, with a particular focus on mouse ESCs.
Figures
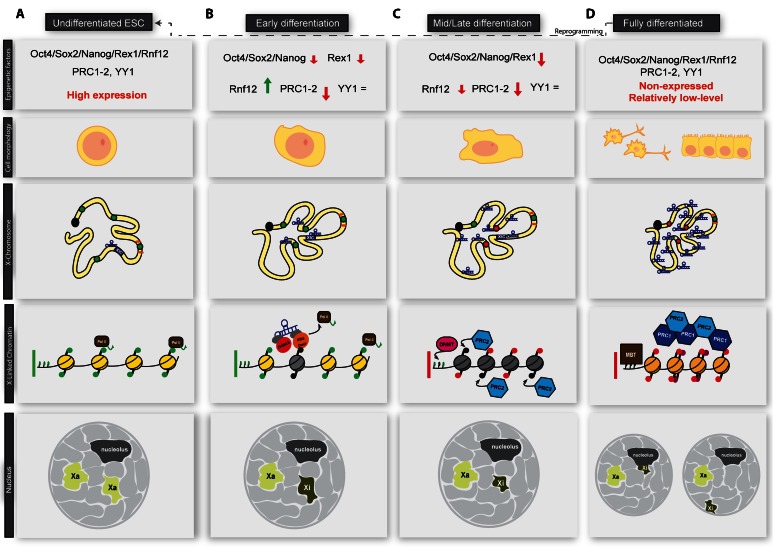
a) In undifferentiated ESCs, chromatin is decompacted and cells are in a fully pluripotent state. Pluripotency factors, master epigenetic regulators (i.e. Polycomb proteins, PRC1/2) are highly expressed and most of the genome is early replicating. b) Early in differentiation, pluripotency factors are downregulated, allowing the monoallelic upregulation of Xist on the future inactive X (Xi). PRC1/2 complexes are also dowregulated [16], YY1 levels remain constant during differentiation [17]. Xist spreads in the 3D neighbourhood and recruits chromatin modifiers like histone deacetylases and/or H3K4me2-3 demethylases to the future inactive X. This recruitment can be either direct or mediated by an adaptor protein. As a consequence of Xist activity, RNA pol II is displaced from actively transcribing promoters. Genes to be silenced start to be relocated inside the Xist-repressive compartment. c) Removal of RNA Pol II from chromatin allows the recruitment of Polycomb proteins (PcGs) and DNA methyltransferases (DNMTs). In particular, the future inactive X becomes enriched for the PRC2 mark H3K27me3 and begins to get compacted as a consequence of gene silencing. Gene relocation is nearly complete at this stage, with only few escapee genes not internalised. CTCFs may serve as a barrier to protect escapee genes. The future inactive X also becomes late replicating. d) PRC2 mark (H3K27me2-3) is in turn recognised by PRC1 and this silencing loop is reinforced by the addition of H2A119ub1* and histone H2A is replaced by the silencing-associated histone variant, macroH2A. Xist spreading is complete at this stage. Chromatin compaction has reached its maximal level and the inactive X translocates to the proximity of the nuclear lamina or the nucleolus. In fully differentiated cells, pluripotency factors are very low. PRC1/2 levels are also usually low. Cell reprogramming can revert the differentiated state to an ESC-like state (iPSC), which is compatible with de novo XCI establishment. *Note: PRC1 can be recruited to the inactive X independently of H3K27me3 mark and it starts to accumulate at low levels on the inactivating X at a similar time [18]
a) In undifferentiated ESCs, chromatin is decompacted and cells are in a fully pluripotent state. Pluripotency factors, master epigenetic regulators (i.e. Polycomb proteins, PRC1/2) are highly expressed and most of the genome is early replicating. b) Early in differentiation, pluripotency factors are downregulated, allowing the monoallelic upregulation of Xist on the future inactive X (Xi). PRC1/2 complexes are also dowregulated [16], YY1 levels remain constant during differentiation [17]. Xist spreads in the 3D neighbourhood and recruits chromatin modifiers like histone deacetylases and/or H3K4me2-3 demethylases to the future inactive X. This recruitment can be either direct or mediated by an adaptor protein. As a consequence of Xist activity, RNA pol II is displaced from actively transcribing promoters. Genes to be silenced start to be relocated inside the Xist-repressive compartment. c) Removal of RNA Pol II from chromatin allows the recruitment of Polycomb proteins (PcGs) and DNA methyltransferases (DNMTs). In particular, the future inactive X becomes enriched for the PRC2 mark H3K27me3 and begins to get compacted as a consequence of gene silencing. Gene relocation is nearly complete at this stage, with only few escapee genes not internalised. CTCFs may serve as a barrier to protect escapee genes. The future inactive X also becomes late replicating. d) PRC2 mark (H3K27me2-3) is in turn recognised by PRC1 and this silencing loop is reinforced by the addition of H2A119ub1* and histone H2A is replaced by the silencing-associated histone variant, macroH2A. Xist spreading is complete at this stage. Chromatin compaction has reached its maximal level and the inactive X translocates to the proximity of the nuclear lamina or the nucleolus. In fully differentiated cells, pluripotency factors are very low. PRC1/2 levels are also usually low. Cell reprogramming can revert the differentiated state to an ESC-like state (iPSC), which is compatible with de novo XCI establishment. *Note: PRC1 can be recruited to the inactive X independently of H3K27me3 mark and it starts to accumulate at low levels on the inactivating X at a similar time [18]
Similar articles
-
The two active X chromosomes in female ESCs block exit from the pluripotent state by modulating the ESC signaling network.Cell Stem Cell. 2014 Feb 6;14(2):203-16. doi: 10.1016/j.stem.2013.11.022. Cell Stem Cell. 2014. PMID: 24506884
-
The Role of Xist in X-Chromosome Dosage Compensation.Trends Cell Biol. 2018 Dec;28(12):999-1013. doi: 10.1016/j.tcb.2018.05.005. Epub 2018 Jun 14. Trends Cell Biol. 2018. PMID: 29910081 Free PMC article. Review.
-
Eutherian mammals use diverse strategies to initiate X-chromosome inactivation during development.Nature. 2011 Apr 21;472(7343):370-4. doi: 10.1038/nature09872. Epub 2011 Apr 6. Nature. 2011. PMID: 21471966
-
Integrated analysis of Xist upregulation and X-chromosome inactivation with single-cell and single-allele resolution.Nat Commun. 2021 Jun 15;12(1):3638. doi: 10.1038/s41467-021-23643-6. Nat Commun. 2021. PMID: 34131144 Free PMC article.
-
Concise review: Pluripotency and the transcriptional inactivation of the female Mammalian X chromosome.Stem Cells. 2012 Jan;30(1):48-54. doi: 10.1002/stem.755. Stem Cells. 2012. PMID: 21997775 Free PMC article. Review.
Cited by
-
Xist condensates: perspectives for therapeutic intervention.Genome Biol. 2025 Jul 21;26(1):215. doi: 10.1186/s13059-025-03666-8. Genome Biol. 2025. PMID: 40691806 Free PMC article. Review.
-
Preventing erosion of X-chromosome inactivation in human embryonic stem cells.Nat Commun. 2022 May 6;13(1):2516. doi: 10.1038/s41467-022-30259-x. Nat Commun. 2022. PMID: 35523820 Free PMC article.
-
Function by Structure: Spotlights on Xist Long Non-coding RNA.Front Mol Biosci. 2017 Dec 19;4:90. doi: 10.3389/fmolb.2017.00090. eCollection 2017. Front Mol Biosci. 2017. PMID: 29302591 Free PMC article. Review.
-
Characterisation of X chromosome status of human extended pluripotent stem cells.Cell Prolif. 2023 May;56(5):e13468. doi: 10.1111/cpr.13468. Epub 2023 May 17. Cell Prolif. 2023. PMID: 37199042 Free PMC article.
-
Long non-coding RNA-polycomb intimate rendezvous.Open Biol. 2020 Sep;10(9):200126. doi: 10.1098/rsob.200126. Epub 2020 Sep 9. Open Biol. 2020. PMID: 32898472 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
