

A draft network of ligand-receptor-mediated multicellular signalling in human
- PMID: 26198319
- PMCID: PMC4525178
- DOI: 10.1038/ncomms8866
A draft network of ligand-receptor-mediated multicellular signalling in human
Erratum in
-
Corrigendum: A draft network of ligand-receptor-mediated multicellular signalling in human.Nat Commun. 2016 Feb 10;7:10706. doi: 10.1038/ncomms10706. Nat Commun. 2016. PMID: 26862012 Free PMC article. No abstract available.
Abstract
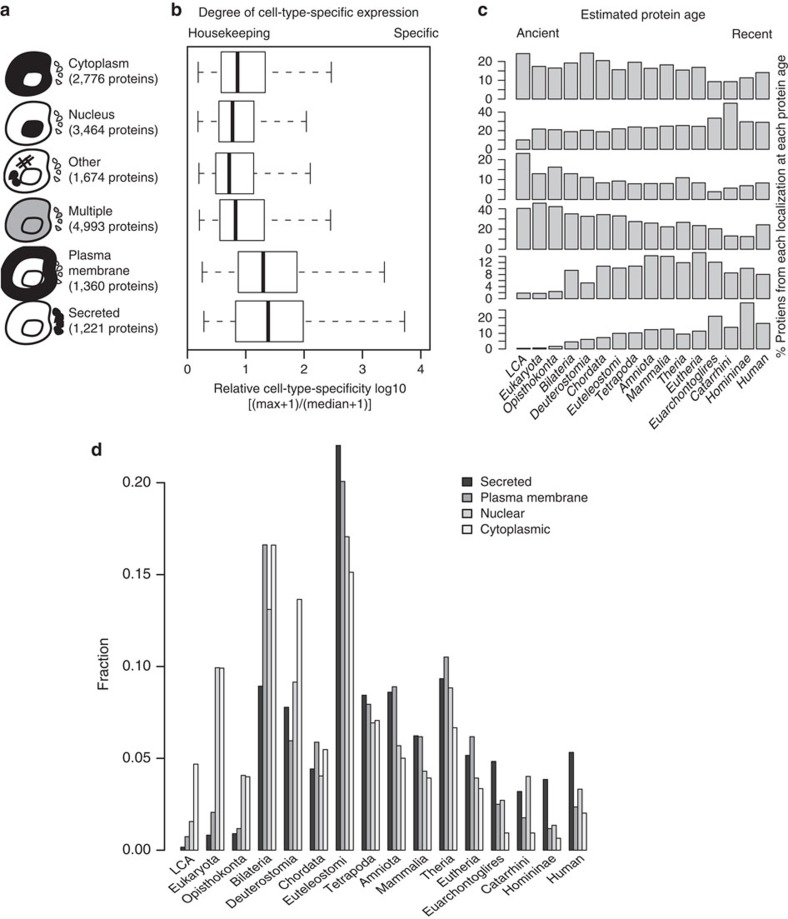
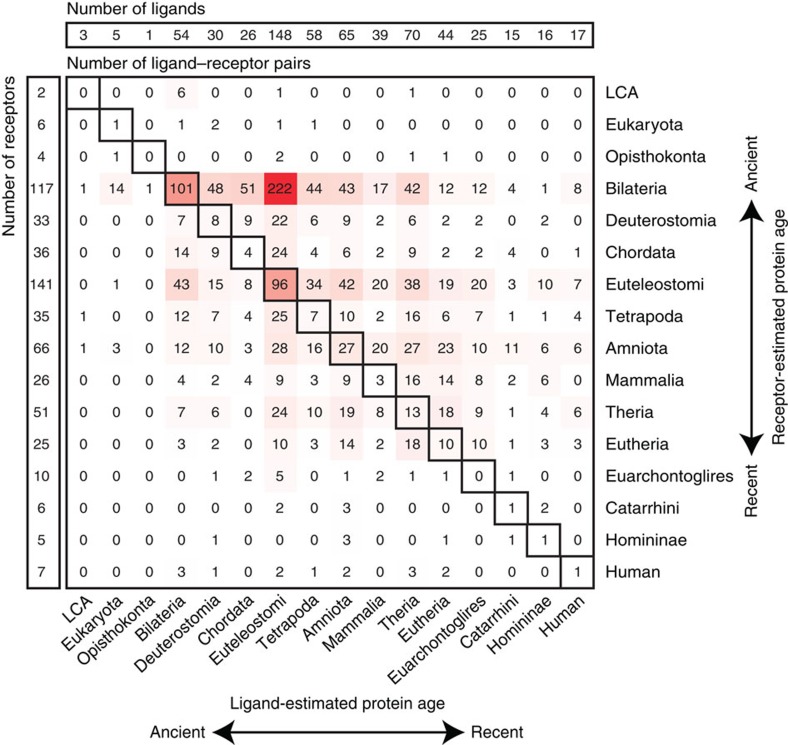
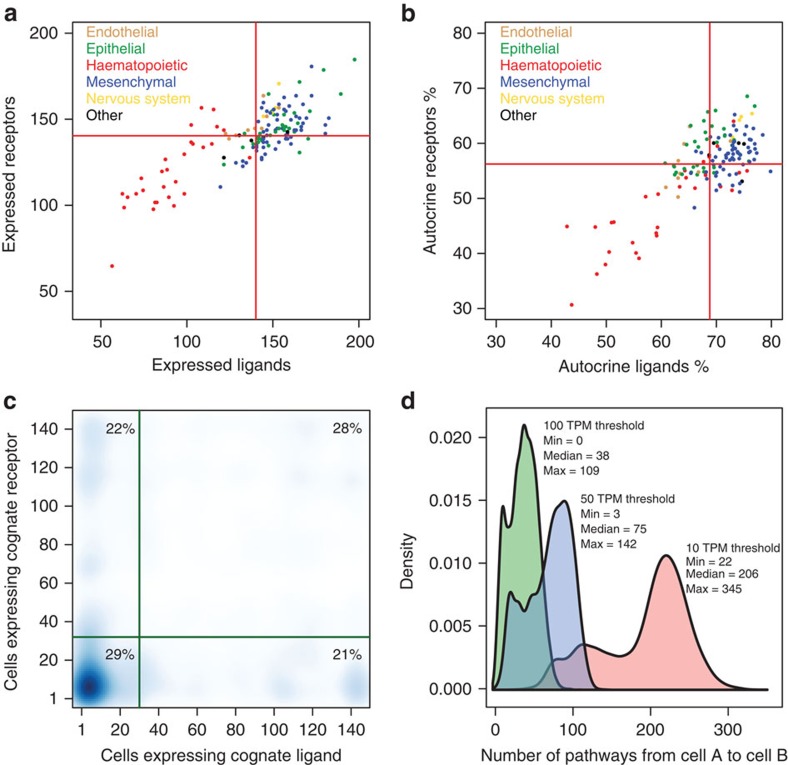
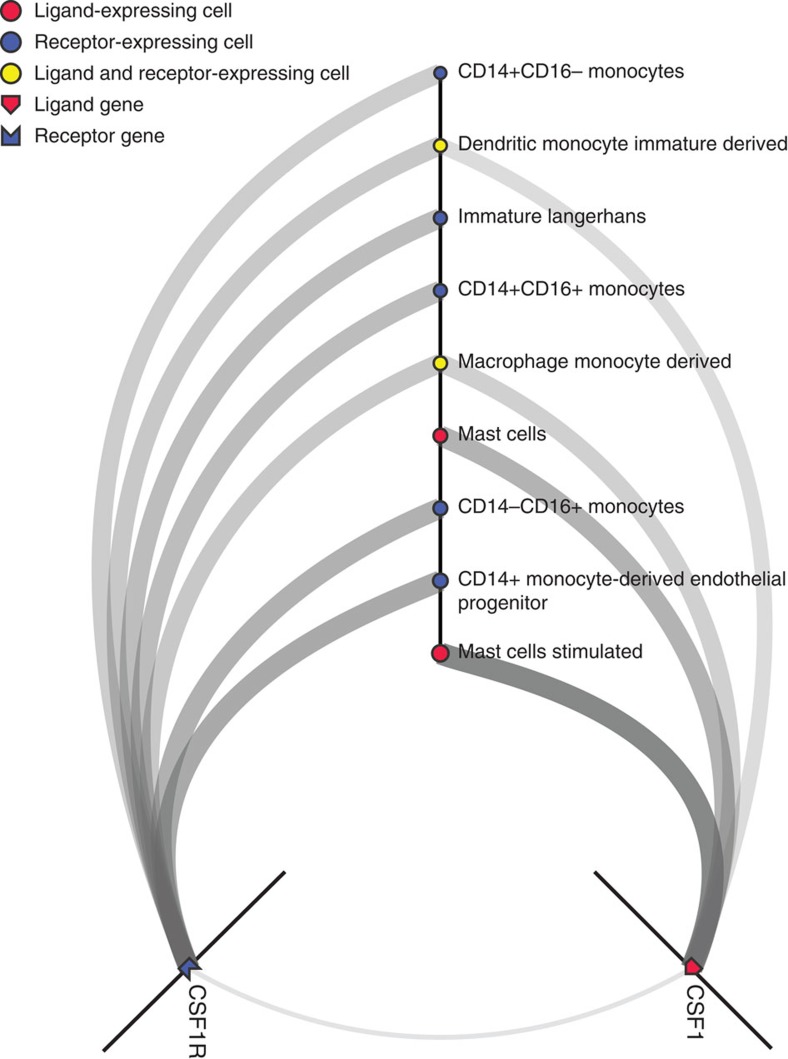
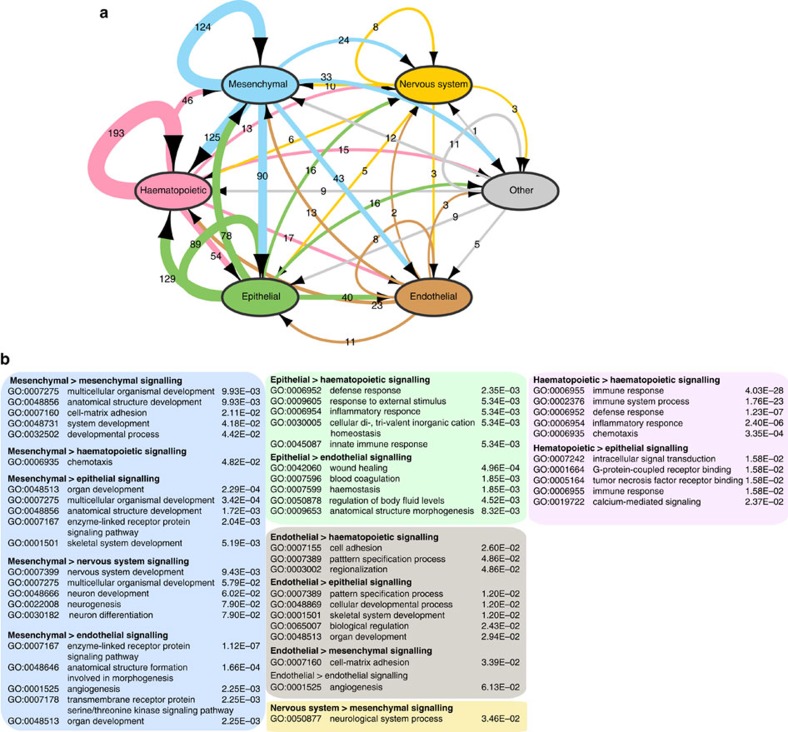
Cell-to-cell communication across multiple cell types and tissues strictly governs proper functioning of metazoans and extensively relies on interactions between secreted ligands and cell-surface receptors. Herein, we present the first large-scale map of cell-to-cell communication between 144 human primary cell types. We reveal that most cells express tens to hundreds of ligands and receptors to create a highly connected signalling network through multiple ligand-receptor paths. We also observe extensive autocrine signalling with approximately two-thirds of partners possibly interacting on the same cell type. We find that plasma membrane and secreted proteins have the highest cell-type specificity, they are evolutionarily younger than intracellular proteins, and that most receptors had evolved before their ligands. We provide an online tool to interactively query and visualize our networks and demonstrate how this tool can reveal novel cell-to-cell interactions with the prediction that mast cells signal to monoblastic lineages via the CSF1-CSF1R interacting pair.
Figures
References
-
- Grosberg R. K. & Strathmann R. R. The evolution of multicellularity: a minor major transition? Annu. Rev. Ecol. Evol. Syst. 38, 621–654 (2007).
-
- Pires-daSilva A. & Sommer R. J. The evolution of signalling pathways in animal development. Nat. Rev. Genet. 4, 39–49 (2003). - PubMed
-
- Gale N. W. et al. Eph receptors and ligands comprise two major specificity subclasses and are reciprocally compartmentalized during embryogenesis. Neuron 17, 9–19 (1996). - PubMed
-
- Kroll K. L. & Amaya E. Transgenic Xenopus embryos from sperm nuclear transplantations reveal FGF signalling requirements during gastrulation. Development 122, 3173–3183 (1996). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
