

Modeling of glucose-induced cAMP oscillations in pancreatic β cells: cAMP rocks when metabolism rolls
- PMID: 26200880
- PMCID: PMC4621809
- DOI: 10.1016/j.bpj.2015.06.024
Modeling of glucose-induced cAMP oscillations in pancreatic β cells: cAMP rocks when metabolism rolls
Abstract
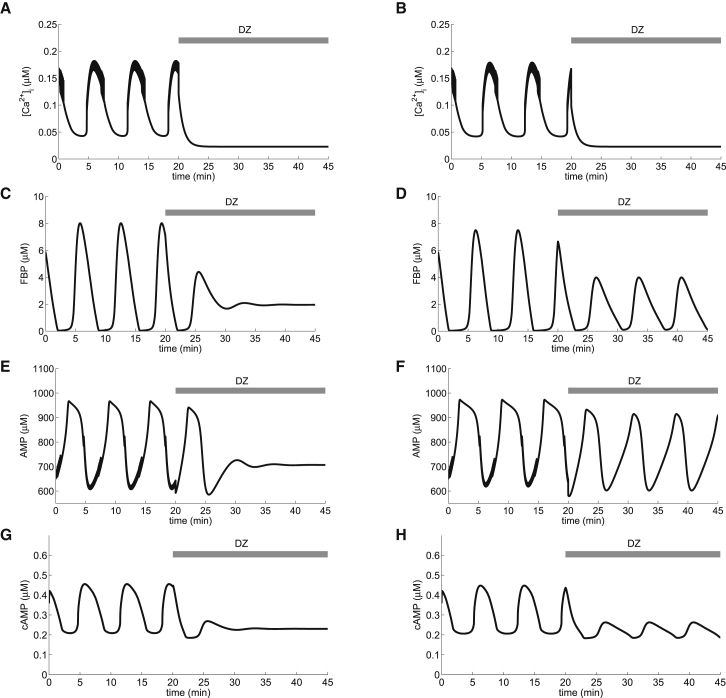
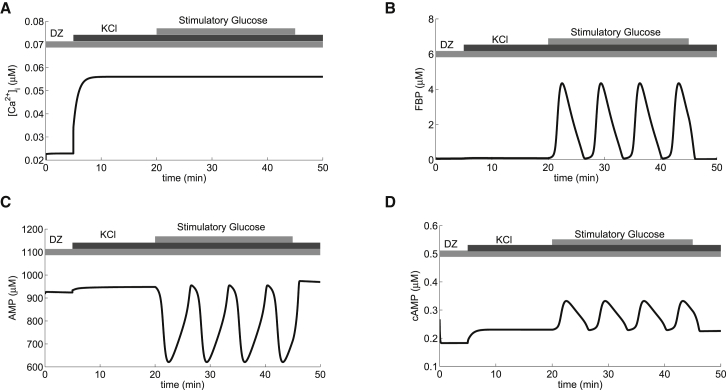
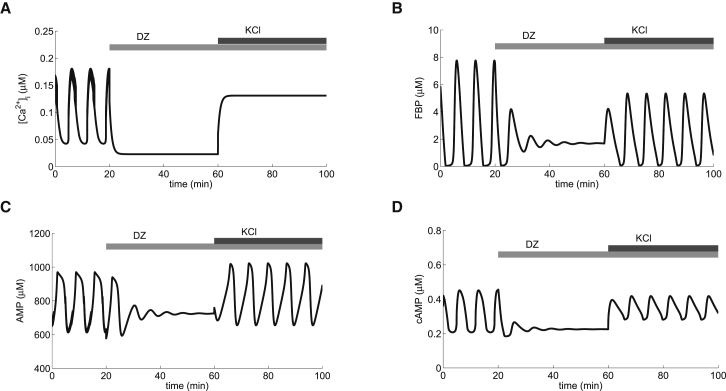
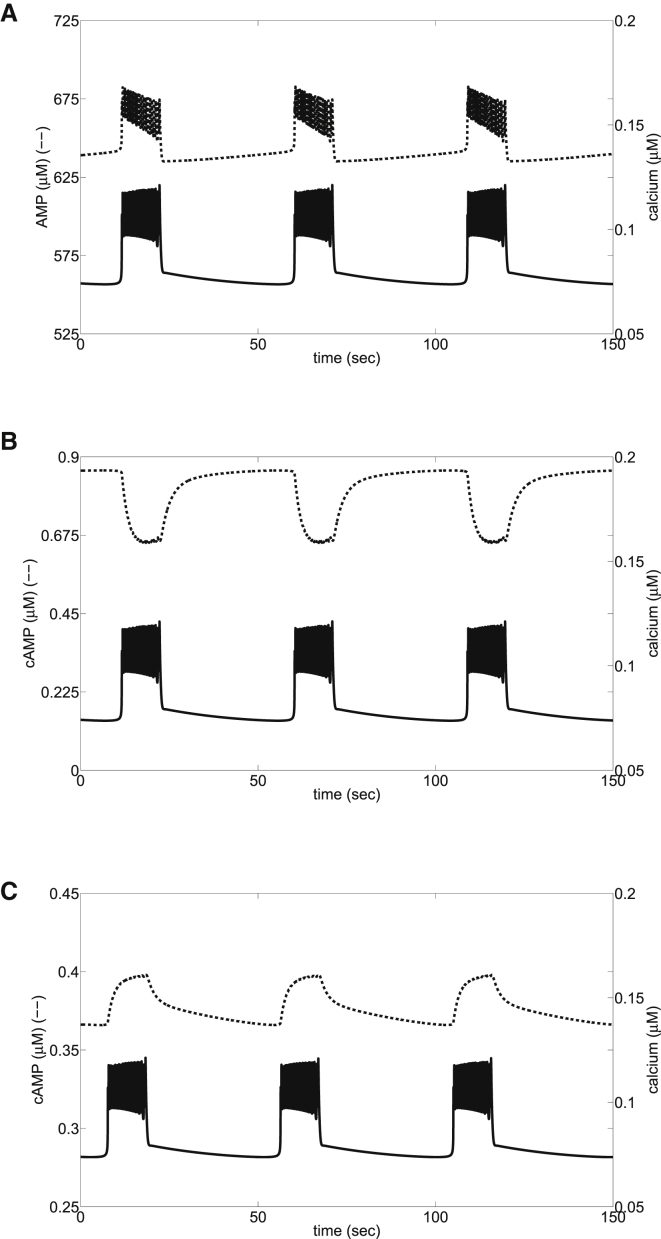
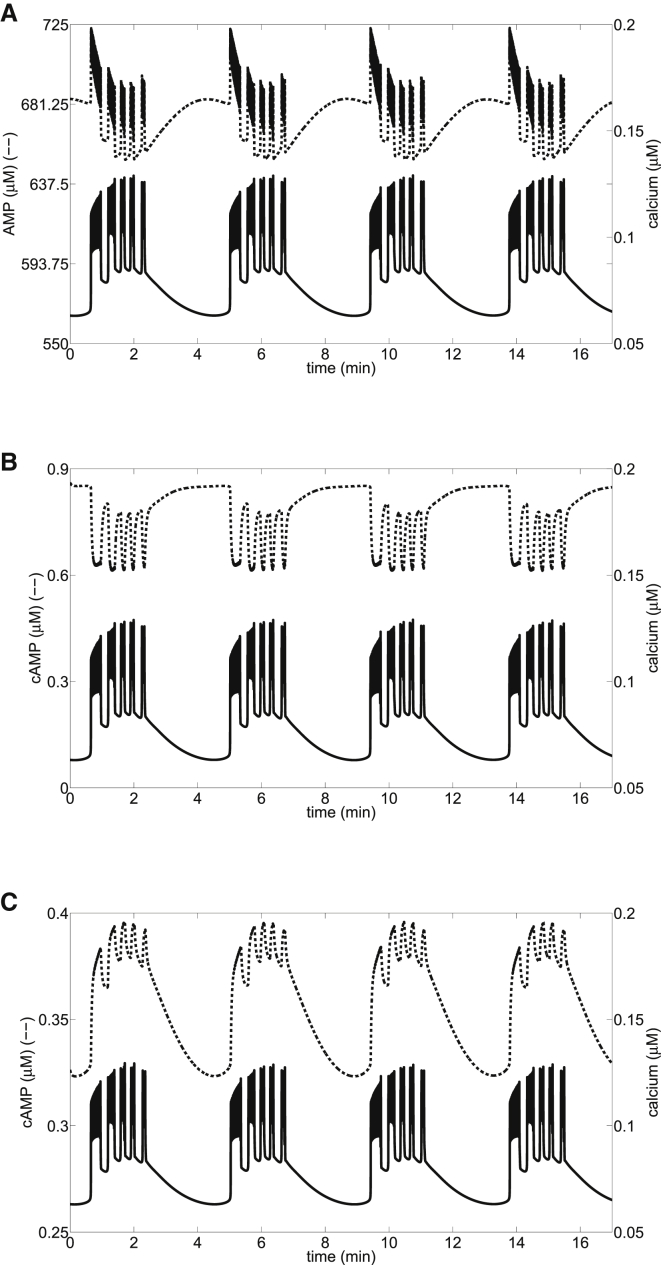
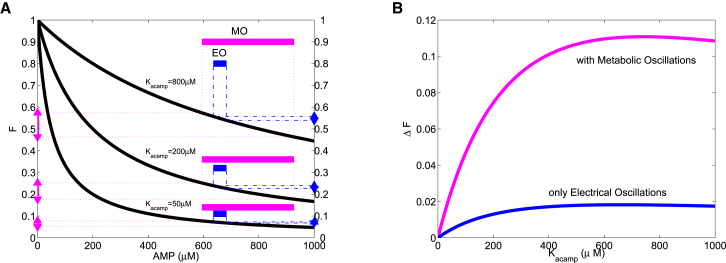
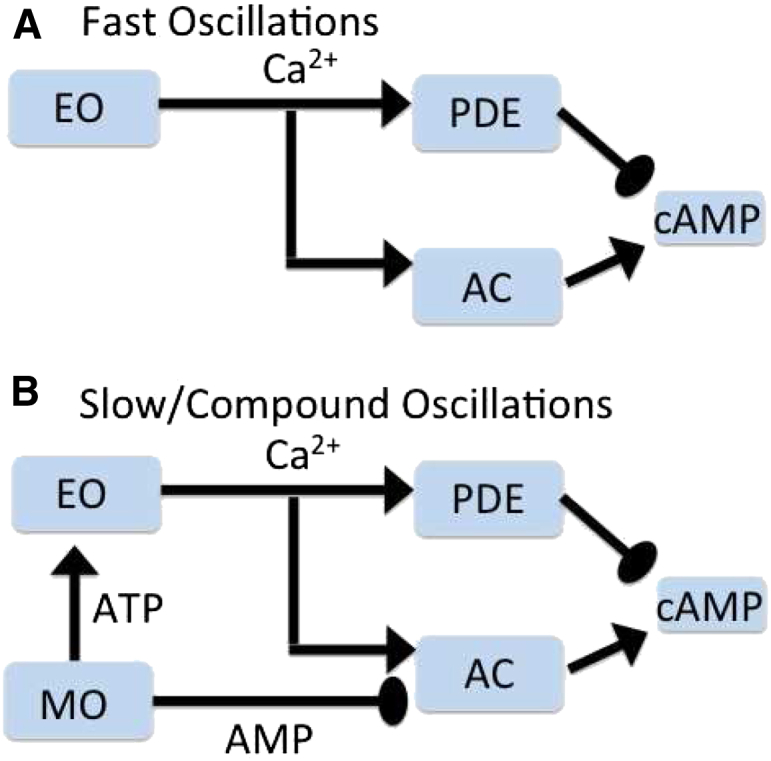
Recent advances in imaging technology have revealed oscillations of cyclic adenosine monophosphate (cAMP) in insulin-secreting cells. These oscillations may be in phase with cytosolic calcium oscillations or out of phase. cAMP oscillations have previously been modeled as driven by oscillations in calcium, based on the known dependence of the enzymes that generate cAMP (adenylyl cyclase) and degrade it (phosphodiesterase). However, cAMP oscillations have also been reported to occur in the absence of calcium oscillations. Motivated by similarities between the properties of cAMP and metabolic oscillations in pancreatic β cells, we propose here that in addition to direct control by calcium, cAMP is controlled by metabolism. Specifically, we hypothesize that AMP inhibits adenylyl cyclase. We incorporate this hypothesis into the dual oscillator model for β cells, in which metabolic (glycolytic) oscillations cooperate with modulation of ion channels and metabolism by calcium. We show that the combination of oscillations in AMP and calcium in the dual oscillator model can account for the diverse oscillatory patterns that have been observed, as well as for experimental perturbations of those patterns. Predictions to further test the model are provided.
Copyright © 2015 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Regulation of cAMP dynamics by Ca2+ and G protein-coupled receptors in the pancreatic beta-cell: a computational approach.Am J Physiol Cell Physiol. 2007 Dec;293(6):C1924-33. doi: 10.1152/ajpcell.00555.2006. Epub 2007 Oct 10. Am J Physiol Cell Physiol. 2007. PMID: 17928534
-
Synchronizing Ca2+ and cAMP oscillations in pancreatic beta-cells: a role for glucose metabolism and GLP-1 receptors? Focus on "regulation of cAMP dynamics by Ca2+ and G protein-coupled receptors in the pancreatic beta-cell: a computational approach".Am J Physiol Cell Physiol. 2008 Jan;294(1):C4-6. doi: 10.1152/ajpcell.00522.2007. Epub 2007 Nov 7. Am J Physiol Cell Physiol. 2008. PMID: 17989206 Free PMC article. No abstract available.
-
Glucose- and hormone-induced cAMP oscillations in α- and β-cells within intact pancreatic islets.Diabetes. 2011 May;60(5):1535-43. doi: 10.2337/db10-1087. Epub 2011 Mar 28. Diabetes. 2011. PMID: 21444924 Free PMC article.
-
cAMP signalling in insulin and glucagon secretion.Diabetes Obes Metab. 2017 Sep;19 Suppl 1:42-53. doi: 10.1111/dom.12993. Diabetes Obes Metab. 2017. PMID: 28466587 Review.
-
Pharmacological modulation of the CO2/HCO3-/pH-, calcium-, and ATP-sensing soluble adenylyl cyclase.Pharmacol Ther. 2018 Oct;190:173-186. doi: 10.1016/j.pharmthera.2018.05.008. Epub 2018 May 26. Pharmacol Ther. 2018. PMID: 29807057 Free PMC article. Review.
Cited by
-
Spatially compartmentalized phase regulation of a Ca2+-cAMP-PKA oscillatory circuit.Elife. 2020 Nov 17;9:e55013. doi: 10.7554/eLife.55013. Elife. 2020. PMID: 33201801 Free PMC article.
-
Glucose-induced cAMP elevation in β-cells involves amplification of constitutive and glucagon-activated GLP-1 receptor signalling.Acta Physiol (Oxf). 2021 Apr;231(4):e13611. doi: 10.1111/apha.13611. Epub 2021 Jan 9. Acta Physiol (Oxf). 2021. PMID: 33369112 Free PMC article.
-
Cyclic AMP links glucose stimulation to somatostatin secretion in δ-cells.J Gen Physiol. 2019 Sep 2;151(9):1062-1065. doi: 10.1085/jgp.201912417. Epub 2019 Aug 14. J Gen Physiol. 2019. PMID: 31413066 Free PMC article.
-
Spatiotemporal orchestration of calcium-cAMP oscillations on AKAP/AC nanodomains is governed by an incoherent feedforward loop.PLoS Comput Biol. 2024 Oct 31;20(10):e1012564. doi: 10.1371/journal.pcbi.1012564. eCollection 2024 Oct. PLoS Comput Biol. 2024. PMID: 39480900 Free PMC article.
-
Ca2+ release or Ca2+ entry, that is the question: what governs Ca2+ oscillations in pancreatic β cells?Am J Physiol Endocrinol Metab. 2023 Jun 1;324(6):E477-E487. doi: 10.1152/ajpendo.00030.2023. Epub 2023 Apr 19. Am J Physiol Endocrinol Metab. 2023. PMID: 37074988 Free PMC article. Review.
References
-
- Dolenšek J., Skelin M., Rupnik M.S. Calcium dependencies of regulated exocytosis in different endocrine cells. Physiol. Res. 2011;60(Suppl 1):S29–S38. - PubMed
-
- Paco S., Margelí M.A., Aguado F. Regulation of exocytotic protein expression and Ca2+-dependent peptide secretion in astrocytes. J. Neurochem. 2009;110:143–156. - PubMed
-
- Braun M., Ramracheya R., Rorsman P. Exocytotic properties of human pancreatic β-cells. Ann. N. Y. Acad. Sci. 2009;1152:187–193. - PubMed
-
- Baggio L.L., Drucker D.J. Biology of incretins: GLP-1 and GIP. Gastroenterology. 2007;132:2131–2157. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
