

Renin expression in developing zebrafish is associated with angiogenesis and requires the Notch pathway and endothelium
- PMID: 26202224
- PMCID: PMC4572395
- DOI: 10.1152/ajprenal.00247.2015
Renin expression in developing zebrafish is associated with angiogenesis and requires the Notch pathway and endothelium
Abstract
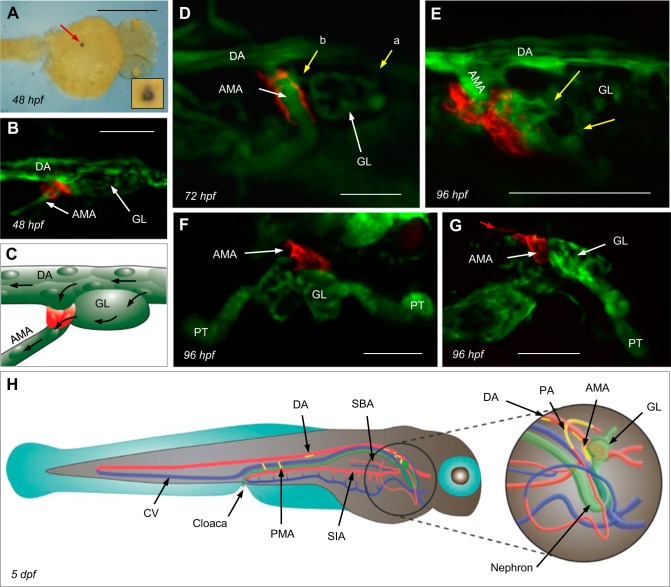
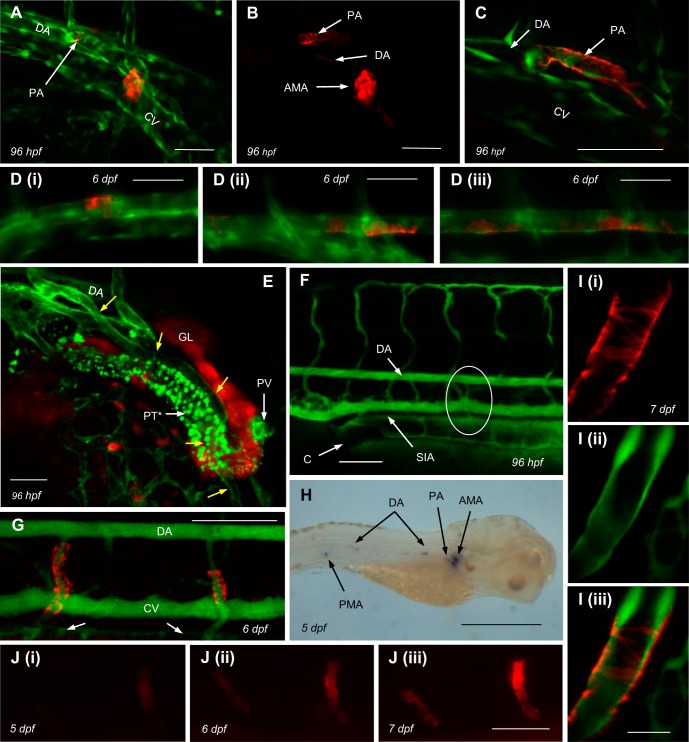
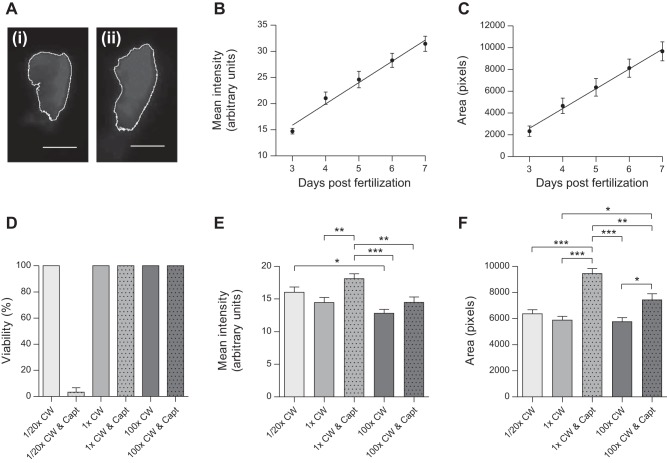
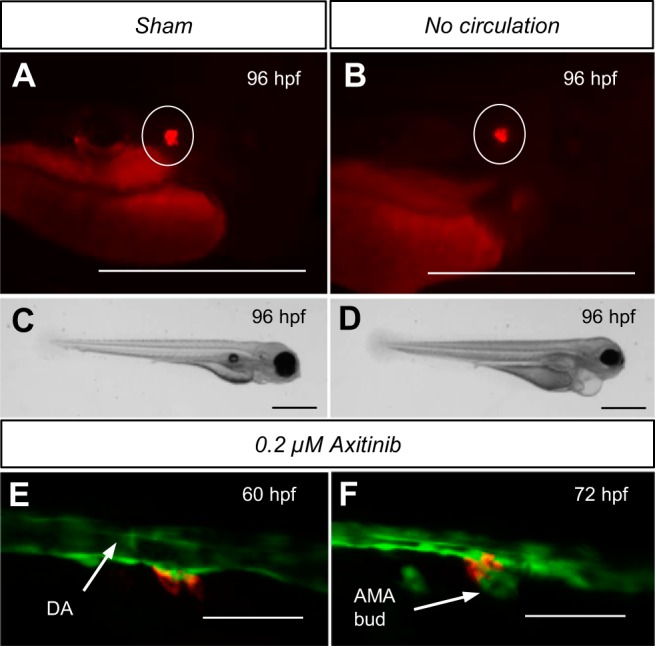
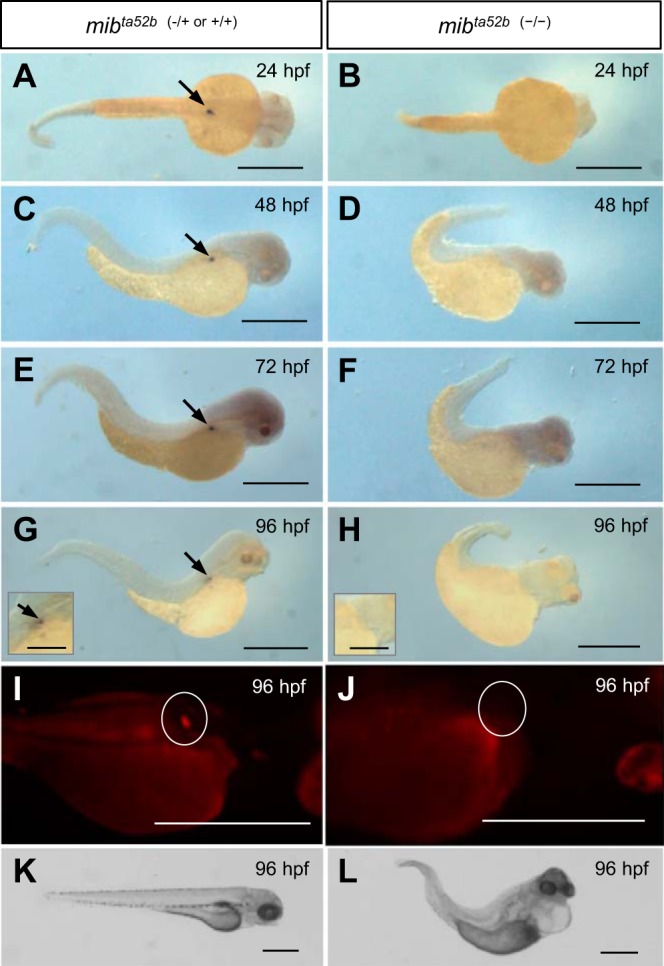
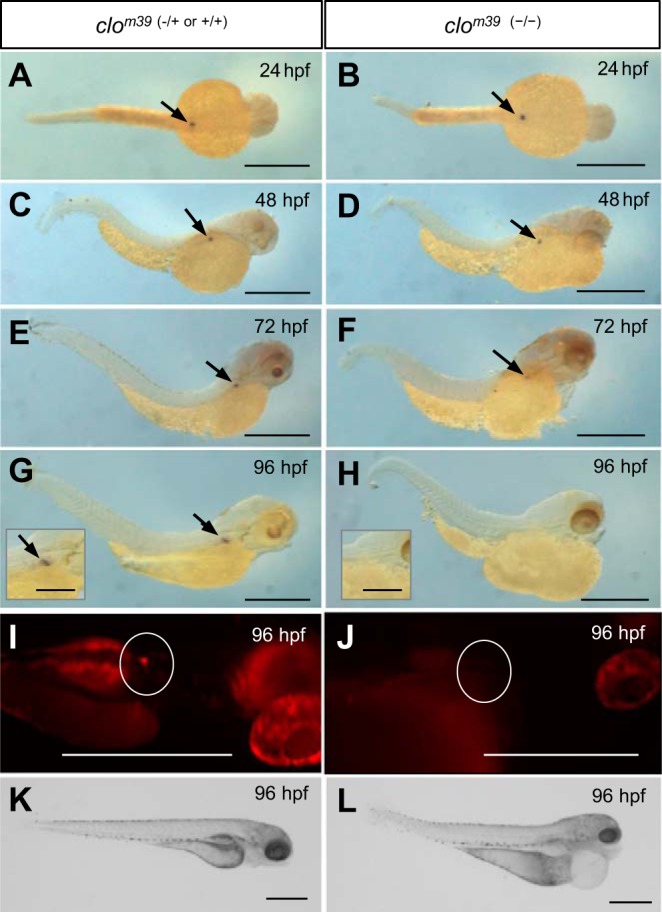
Although renin is a critical regulatory enzyme of the cardiovascular system, its roles in organogenesis and the establishment of cardiovascular homeostasis remain unclear. Mammalian renin-expressing cells are widespread in embryonic kidneys but are highly restricted, specialized endocrine cells in adults. With a functional pronephros, embryonic zebrafish are ideal for delineating the developmental functions of renin-expressing cells and the mechanisms governing renin transcription. Larval zebrafish renin expression originates in the mural cells of the juxtaglomerular anterior mesenteric artery and subsequently at extrarenal sites. The role of renin was determined by assessing responses to renin-angiotensin system blockade, salinity variation, and renal perfusion ablation. Renin expression did not respond to renal flow ablation but was modulated by inhibition of angiotensin-converting enzyme and altered salinity. Our data in larval fish are consistent with conservation of renin's physiological functions. Using transgenic renin reporter fish, with mindbomb and cloche mutants, we show that Notch signaling and the endothelium are essential for developmental renin expression. After inhibition of angiogenesis, renin-expressing cells precede angiogenic sprouts. Arising from separate lineages, but relying on mutual interplay with endothelial cells, renin-expressing cells are among the earliest mural cells observed in larval fish, performing both endocrine and paracrine functions.
Keywords: angiogenesis; endothelium; notch; renin; zebrafish.
Copyright © 2015 the American Physiological Society.
Figures
Similar articles
-
Notch signalling limits angiogenic cell behaviour in developing zebrafish arteries.Nature. 2007 Feb 15;445(7129):781-4. doi: 10.1038/nature05577. Epub 2007 Jan 28. Nature. 2007. PMID: 17259972
-
carbamoyl-phosphate synthetase 2, aspartate transcarbamylase, and dihydroorotase (cad) regulates Notch signaling and vascular development in zebrafish.Dev Dyn. 2015 Jan;244(1):1-9. doi: 10.1002/dvdy.24209. Epub 2014 Nov 17. Dev Dyn. 2015. PMID: 25294789
-
UXT potentiates angiogenesis by attenuating Notch signaling.Development. 2015 Feb 15;142(4):774-86. doi: 10.1242/dev.112532. Epub 2015 Jan 23. Development. 2015. PMID: 25617435 Free PMC article.
-
Notch signalling in ischaemia-induced angiogenesis.Biochem Soc Trans. 2009 Dec;37(Pt 6):1221-7. doi: 10.1042/BST0371221. Biochem Soc Trans. 2009. PMID: 19909251 Free PMC article. Review.
-
Notch function in the vasculature: insights from zebrafish, mouse and man.Bioessays. 2004 Mar;26(3):225-34. doi: 10.1002/bies.20004. Bioessays. 2004. PMID: 14988924 Review.
Cited by
-
Juxtaglomerular Cell Phenotypic Plasticity.High Blood Press Cardiovasc Prev. 2017 Sep;24(3):231-242. doi: 10.1007/s40292-017-0212-5. Epub 2017 May 19. High Blood Press Cardiovasc Prev. 2017. PMID: 28527017 Free PMC article. Review.
-
Investigating the RAS can be a fishy business: interdisciplinary opportunities using Zebrafish.Clin Sci (Lond). 2018 Dec 5;132(23):2469-2481. doi: 10.1042/CS20180721. Print 2018 Dec 12. Clin Sci (Lond). 2018. PMID: 30518571 Free PMC article. Review.
-
Angiotensin II Signal Transduction: An Update on Mechanisms of Physiology and Pathophysiology.Physiol Rev. 2018 Jul 1;98(3):1627-1738. doi: 10.1152/physrev.00038.2017. Physiol Rev. 2018. PMID: 29873596 Free PMC article. Review.
-
Small fish, big discoveries: zebrafish shed light on microbial biomarkers for neuro-immune-cardiovascular health.Front Physiol. 2023 Jun 1;14:1186645. doi: 10.3389/fphys.2023.1186645. eCollection 2023. Front Physiol. 2023. PMID: 37324381 Free PMC article. Review.
-
Basement membrane defects in CD151-associated glomerular disease.Pediatr Nephrol. 2022 Dec;37(12):3105-3115. doi: 10.1007/s00467-022-05447-y. Epub 2022 Mar 12. Pediatr Nephrol. 2022. PMID: 35278129 Free PMC article.
References
-
- Bollig F, Mehringer R, Perner B, Hartung C, Schafer M, Schartl M, Volff JN, Winkler C, Englert C. Identification and comparative expression analysis of a second wt1 gene in zebrafish. Dev Dyn 235: 554–561, 2006. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases