

doi: 10.1128/JVI.01604-15.
Epub 2015 Jul 22.
The Epstein-Barr Virus BDLF4 Gene Is Required for Efficient Expression of Viral Late Lytic Genes
Affiliations
- PMID: 26202235
- PMCID: PMC4577904
- DOI: 10.1128/JVI.01604-15
Item in Clipboard
The Epstein-Barr Virus BDLF4 Gene Is Required for Efficient Expression of Viral Late Lytic Genes
J Virol.
2015 Oct.
Abstract
Epstein-Barr virus (EBV) is a gammaherpesvirus, associated with infectious mononucleosis and various types of malignancy. We focused here on the BDLF4 gene of EBV and identified it as a lytic gene, expressed with early kinetics. Viral late gene expression of the BDLF4 knockout strain was severely restricted; this could be restored by an exogenous supply of BDLF4. These results indicate that BDLF4 is important for the EBV lytic replication cycle, especially in late gene expression.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
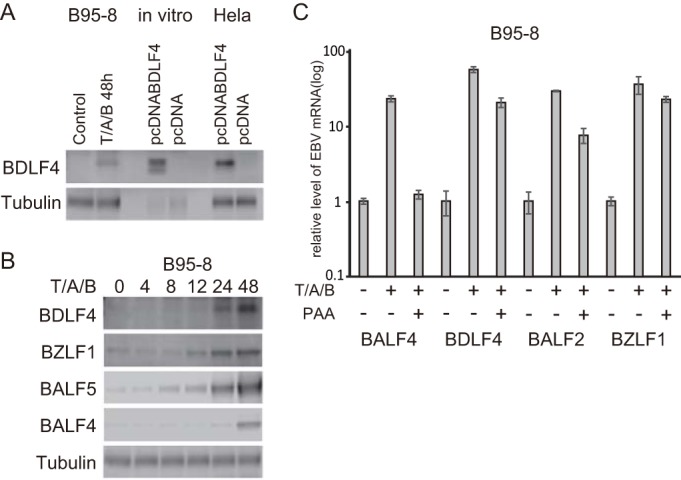
Identification of BDLF4 protein. (A) B95-8 cell lysate, BDLF4 protein synthesized in vitro, and BDLF4 expressed in HeLa cells were subjected to Western blotting. B95-8 cells were incubated with TPA (200 ng/ml), A23187 (0.5 μM), and sodium butyrate (5 mM) (T/A/B) to induce the lytic cycle. (B) Expression levels of BDLF4, BZLF1, BALF5 (Pol), and BALF4 (gB) in B95-8 cells were assessed by Western blotting at the indicated hours after lytic induction. (C) B95-8 cells were treated with PAA to examine the kinetics of BDLF4 expression. Lytic induction was carried out for 48 h (T/A/B) in the presence or absence of 400 μg/ml of PAA. Cells lysates were subjected to RT-PCR using BALF4 (gB), BDLF4, BALF2, and BZLF1 primers as described previously (24).
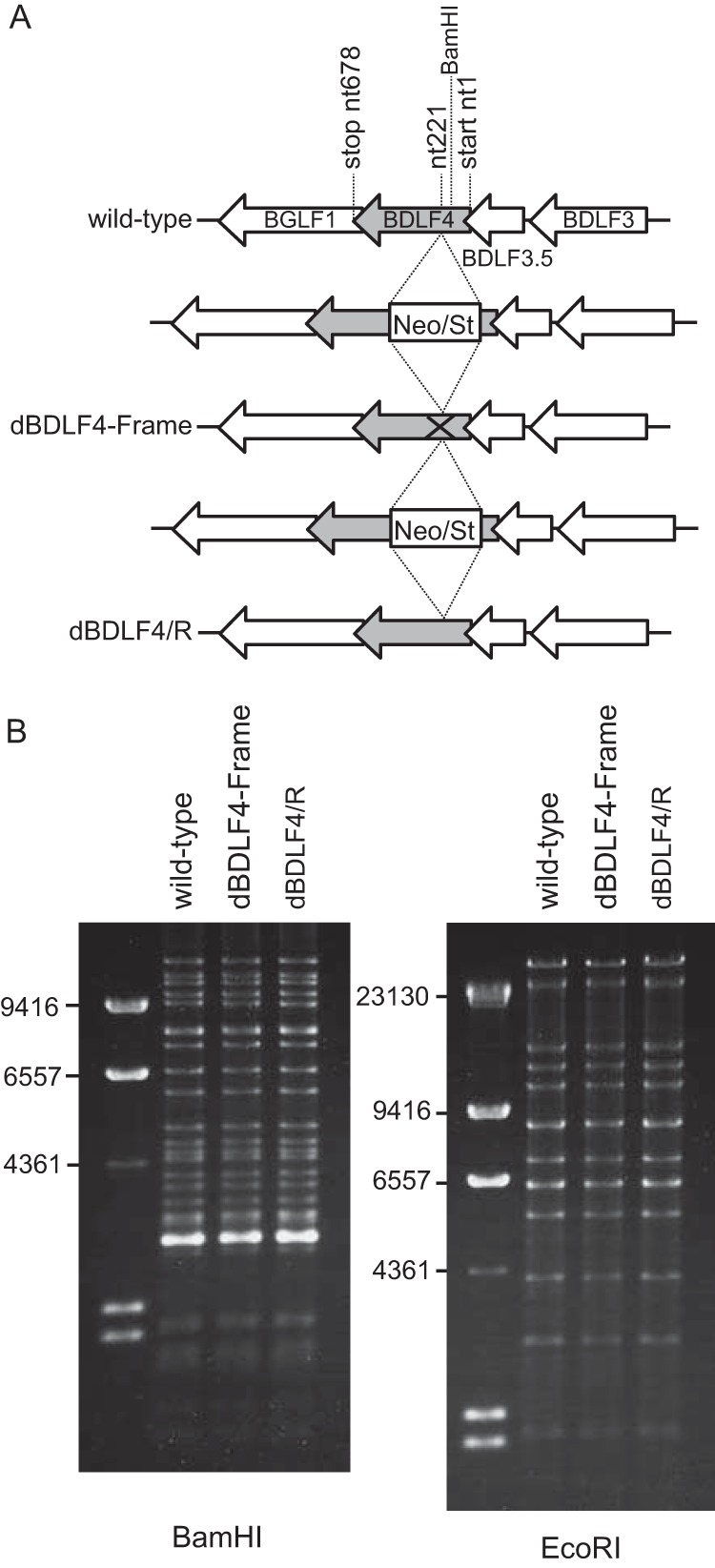
Construction of a point mutant of EBV BDLF4. (A) BDLF4 knockout EBV and its revertant virus were generated using a BAC system in Escherichia coli (25, 26). The neomycin resistance and streptomycin sensitivity genes (Neo/St) were inserted between nucleotides 220 (nt220) and nt221 of the BDLF4 gene. The Neo/St cassette was then replaced by a BDLF4 sequence with a one-nucleotide deletion at T (nt222) (dBDLF4-Frame). This one nucleotide excision caused frame shifting in the BDLF4 gene at residue 74 and creation of a new termination signal after synthesizing 12 nonsense, unrelated amino acids. The Neo/St cassette was again inserted and replaced with a wild-type BDLF4 sequence to make the revertant virus dBDLF4/R. (B) Electrophoresis of the recombinant viruses. EBV BAC DNAs were digested with BamHI or EcoRI and separated in an agarose gel.
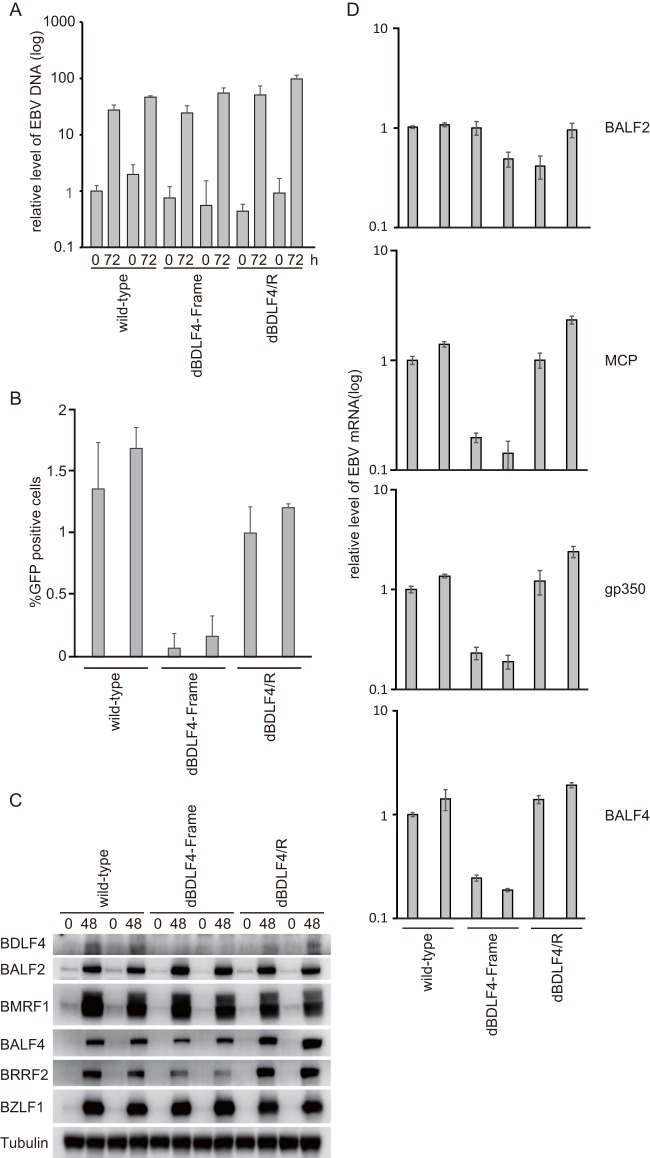
DNA synthesis (A), progeny production (B), viral protein expression (C), and viral mRNA levels (D) in wild-type, BDLF4 mutant, and revertant viruses. (A) Viral DNA was prepared from HEK293 cells that carry the indicated recombinant EBV-BAC genomes at 0 or 72 h after transfection of the BZLF1 expression vector and was analyzed by quantitative real-time PCR. (B) Supernatants were collected at 72 h after transfection of pcDNA-BZLF1. Virus titers in 1 ml of supernatants were determined by examining the levels of green fluorescent protein (GFP) expression in Akata(-) cells after 2 days by fluorescence-activated cell sorter (FACS) analysis. (C) HEK293 cells were induced lytically by transfection of pcDNA-BZLF1. Whole-cell lysates were prepared at 0 and 48 h and analyzed by Western blotting using anti-BDLF4, anti-BALF2, anti-BMRF1, anti-BALF4 (gB), anti-BRRF2, anti-BZLF1, and anti-tubulin antibodies. (D) Real-time RT-PCR was carried out to determine expression of viral late genes (MCP [major capsid protein], gP350, and BALF4 [gB]) and an early gene. HEK293 cells carrying the indicated EBV genomes were transfected with pcDNA-BZLF1 and harvested after 72 h. As shown in this figure, two independent cell clones were tested for wild-type, dBDLF4-Frame, and dBDLF4/R. Bars represent the means and standard deviations of results from three independent experiments.
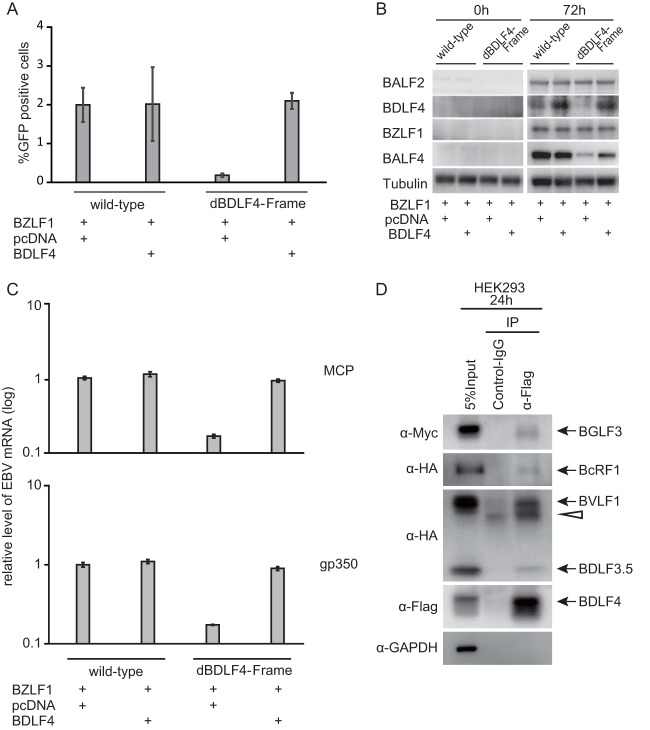
Loss of BDLF4 is responsible for the inhibition of late gene expression (A) HEK293 cells latently infected with EBV were transfected with the indicated expression vector and harvested after 96 h. After freeze-thawing and centrifugation, the supernatants were cultured with Akata(-) cells. FACS analysis was performed to count GFP-positive cells. (B) Levels of viral proteins. HEK293 cells with recombinant EBV were transfected with the indicated vector. Whole-cell lysates were prepared at 0 and 72 h and analyzed by Western blotting with anti-BALF2, anti-BDLF4, anti-BZLF1, anti-BALF4 (gB), and anti-tubulin. (C) Levels of viral late mRNAs. HEK293 cells with recombinant EBVs were transfected with the indicated vector. The mRNAs were prepared at 72 h and analyzed by real-time RT-PCR. (D) Association of BDLF4 with other late gene regulators. HEK293 cells were transfected with expression vectors for Flag-BDLF4, Myc-BGLF3, hemagglutinin-BcRF1 (HA-BcRF1), HA-BVLF1, and HA-BDLF3.5. Immunoprecipitation (IP) was carried out using anti-Flag antibody, and the results were detected by Western blotting using anti-Myc, anti-HA, and anti-Flag antibodies. Anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as a negative control. The arrowhead indicates immunoglobulin.
Similar articles
-
Epstein-Barr Virus BKRF4 Gene Product Is Required for Efficient Progeny Production.J Virol. 2017 Nov 14;91(23):e00975-17. doi: 10.1128/JVI.00975-17. Print 2017 Dec 1. J Virol. 2017. PMID: 28904200 Free PMC article.
-
S-Like-Phase Cyclin-Dependent Kinases Stabilize the Epstein-Barr Virus BDLF4 Protein To Temporally Control Late Gene Transcription.J Virol. 2019 Apr 3;93(8):e01707-18. doi: 10.1128/JVI.01707-18. Print 2019 Apr 15. J Virol. 2019. PMID: 30700607 Free PMC article.
-
Roles of Epstein-Barr virus BGLF3.5 gene and two upstream open reading frames in lytic viral replication in HEK293 cells.Virology. 2015 Sep;483:44-53. doi: 10.1016/j.virol.2015.04.007. Epub 2015 May 15. Virology. 2015. PMID: 25965794
-
Regulation and dysregulation of Epstein-Barr virus latency: implications for the development of autoimmune diseases.Autoimmunity. 2008 May;41(4):298-328. doi: 10.1080/08916930802024772. Autoimmunity. 2008. PMID: 18432410 Review.
-
Lytic cycle switches of oncogenic human gammaherpesviruses.Adv Cancer Res. 2007;97:81-109. doi: 10.1016/S0065-230X(06)97004-3. Adv Cancer Res. 2007. PMID: 17419942 Review.
Cited by
-
Herpesvirus Late Gene Expression: A Viral-Specific Pre-initiation Complex Is Key.Front Microbiol. 2016 Jun 6;7:869. doi: 10.3389/fmicb.2016.00869. eCollection 2016. Front Microbiol. 2016. PMID: 27375590 Free PMC article. Review.
-
Lytic Replication and Reactivation from B Cells Is Not Required for Establishing or Maintaining Gammaherpesvirus Latency In Vivo.J Virol. 2022 Jun 22;96(12):e0069022. doi: 10.1128/jvi.00690-22. Epub 2022 Jun 1. J Virol. 2022. PMID: 35647668 Free PMC article.
-
Comparative Analysis of the Humoral Immune Response to the EBV Proteome across EBV-Related Malignancies.Cancer Epidemiol Biomarkers Prev. 2023 May 1;32(5):687-696. doi: 10.1158/1055-9965.EPI-22-0452. Cancer Epidemiol Biomarkers Prev. 2023. PMID: 36788424 Free PMC article.
-
Antitumor activity of cyclin-dependent kinase inhibitor alsterpaullone in Epstein-Barr virus-associated lymphoproliferative disorders.Cancer Sci. 2020 Jan;111(1):279-287. doi: 10.1111/cas.14241. Epub 2019 Dec 11. Cancer Sci. 2020. PMID: 31743514 Free PMC article.
-
The C-Terminus of Epstein-Barr Virus BRRF2 Is Required for its Proper Localization and Efficient Virus Production.Front Microbiol. 2017 Jan 31;8:125. doi: 10.3389/fmicb.2017.00125. eCollection 2017. Front Microbiol. 2017. PMID: 28197146 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
