

Redundant Function of Plasmacytoid and Conventional Dendritic Cells Is Required To Survive a Natural Virus Infection
- PMID: 26202250
- PMCID: PMC4577909
- DOI: 10.1128/JVI.01024-15
Redundant Function of Plasmacytoid and Conventional Dendritic Cells Is Required To Survive a Natural Virus Infection
Abstract
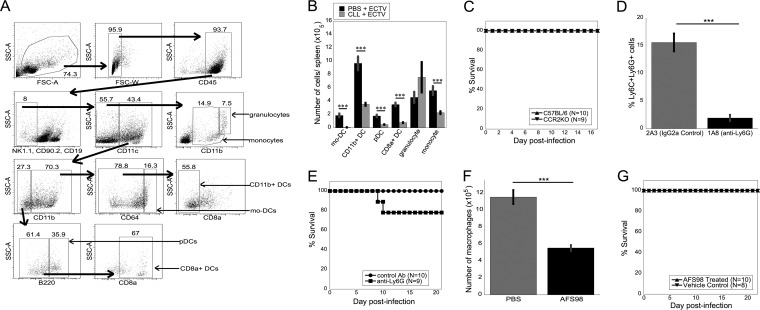
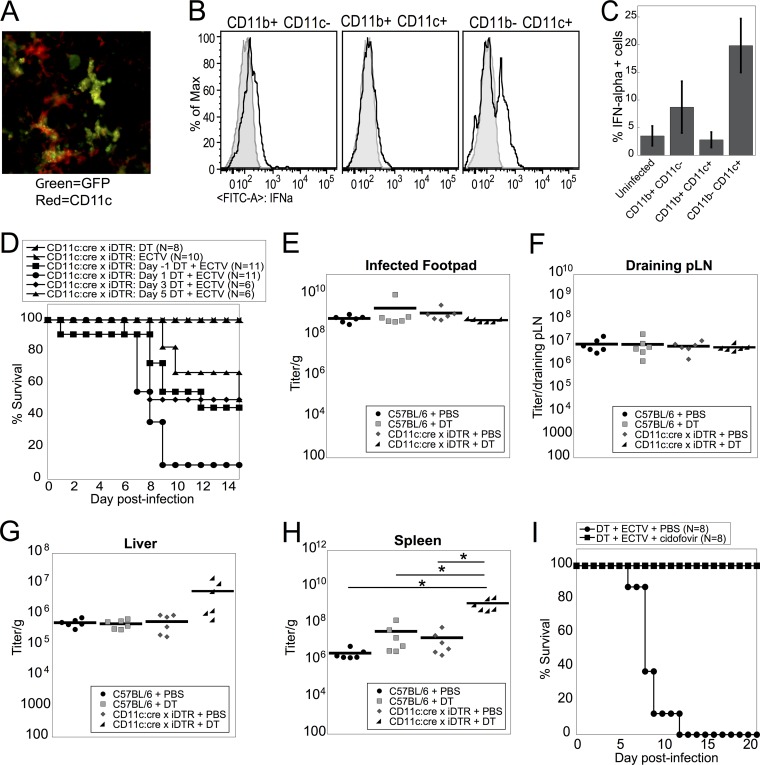
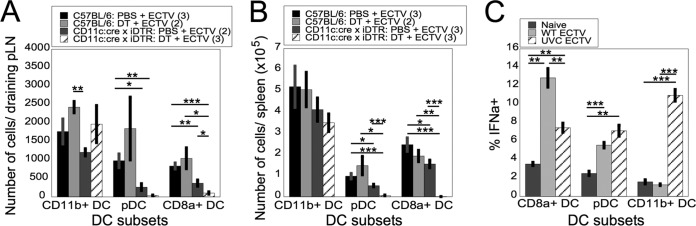
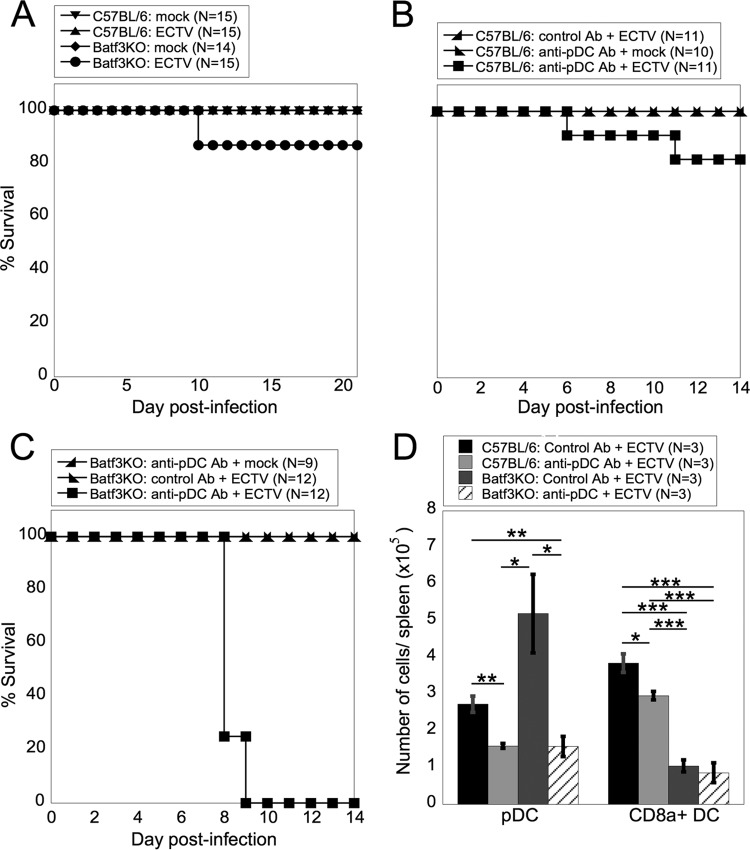
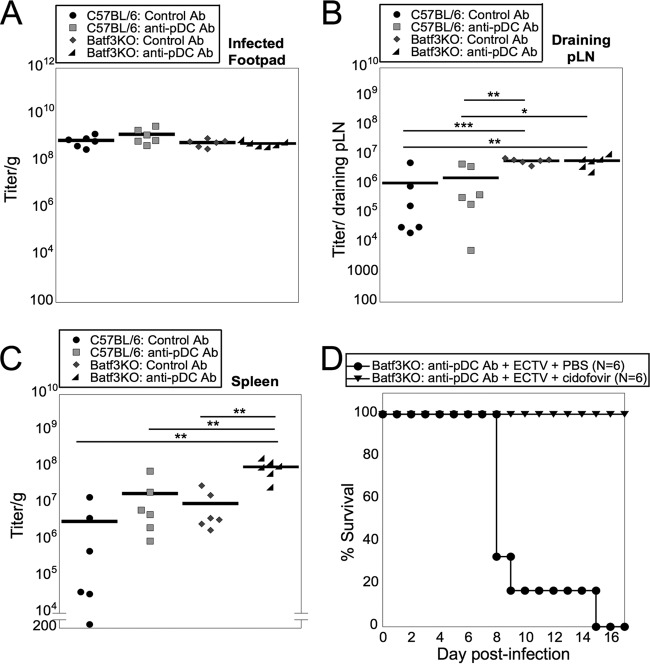
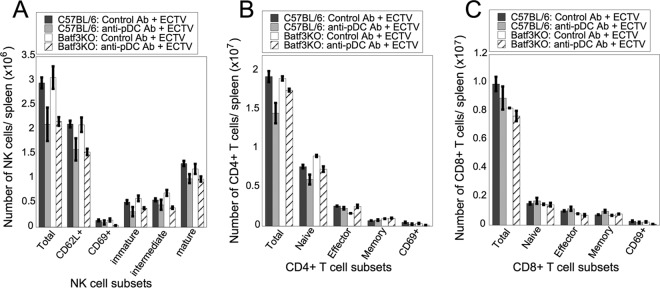
Viruses that spread systemically from a peripheral site of infection cause morbidity and mortality in the human population. Innate myeloid cells, including monocytes, macrophages, monocyte-derived dendritic cells (mo-DC), and dendritic cells (DC), respond early during viral infection to control viral replication, reducing virus spread from the peripheral site. Ectromelia virus (ECTV), an orthopoxvirus that naturally infects the mouse, spreads systemically from the peripheral site of infection and results in death of susceptible mice. While phagocytic cells have a requisite role in the response to ECTV, the requirement for individual myeloid cell populations during acute immune responses to peripheral viral infection is unclear. In this study, a variety of myeloid-specific depletion methods were used to dissect the roles of individual myeloid cell subsets in the survival of ECTV infection. We showed that DC are the primary producers of type I interferons (T1-IFN), requisite cytokines for survival, following ECTV infection. DC, but not macrophages, monocytes, or granulocytes, were required for control of the virus and survival of mice following ECTV infection. Depletion of either plasmacytoid DC (pDC) alone or the lymphoid-resident DC subset (CD8α(+) DC) alone did not confer lethal susceptibility to ECTV. However, the function of at least one of the pDC or CD8α(+) DC subsets is required for survival of ECTV infection, as mice depleted of both populations were susceptible to ECTV challenge. The presence of at least one of these DC subsets is sufficient for cytokine production that reduces ECTV replication and virus spread, facilitating survival following infection.
Importance: Prior to the eradication of variola virus, the orthopoxvirus that causes smallpox, one-third of infected people succumbed to the disease. Following successful eradication of smallpox, vaccination rates with the smallpox vaccine have significantly dropped. There is now an increasing incidence of zoonotic orthopoxvirus infections for which there are no effective treatments. Moreover, the safety of the smallpox vaccine is of great concern, as complications may arise, resulting in morbidity. Like many viruses that cause significant human diseases, orthopoxviruses spread from a peripheral site of infection to become systemic. This study elucidates the early requirement for innate immune cells in controlling a peripheral infection with ECTV, the causative agent of mousepox. We report that there is redundancy in the function of two innate immune cell subsets in controlling virus spread early during infection. The viral control mediated by these cell subsets presents a potential target for therapies and rational vaccine design.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Deficiency in Th2 cytokine responses exacerbate orthopoxvirus infection.PLoS One. 2015 Mar 9;10(3):e0118685. doi: 10.1371/journal.pone.0118685. eCollection 2015. PLoS One. 2015. PMID: 25751266 Free PMC article.
-
The Pathogenesis and Immunobiology of Mousepox.Adv Immunol. 2016;129:251-76. doi: 10.1016/bs.ai.2015.10.001. Epub 2015 Nov 21. Adv Immunol. 2016. PMID: 26791861 Review.
-
Loss of Resistance to Mousepox during Chronic Lymphocytic Choriomeningitis Virus Infection Is Associated with Impaired T-Cell Responses and Can Be Rescued by Immunization.J Virol. 2020 Feb 14;94(5):e01832-19. doi: 10.1128/JVI.01832-19. Print 2020 Feb 14. J Virol. 2020. PMID: 31826990 Free PMC article.
-
Ectromelia virus: the causative agent of mousepox.J Gen Virol. 2005 Oct;86(Pt 10):2645-2659. doi: 10.1099/vir.0.81090-0. J Gen Virol. 2005. PMID: 16186218 Review.
-
Dendritic cells during mousepox: The role of delayed apoptosis in the pathogenesis of infection.Microb Pathog. 2017 Aug;109:99-109. doi: 10.1016/j.micpath.2017.05.037. Epub 2017 May 26. Microb Pathog. 2017. PMID: 28554653
Cited by
-
Synoviocyte-targeted therapy synergizes with TNF inhibition in arthritis reversal.Sci Adv. 2020 Jun 26;6(26):eaba4353. doi: 10.1126/sciadv.aba4353. eCollection 2020 Jun. Sci Adv. 2020. PMID: 32637608 Free PMC article.
-
Langerhans Cells Orchestrate the Protective Antiviral Innate Immune Response in the Lymph Node.Cell Rep. 2019 Dec 3;29(10):3047-3059.e3. doi: 10.1016/j.celrep.2019.10.118. Cell Rep. 2019. PMID: 31801072 Free PMC article.
-
Resident plasmacytoid dendritic cells patrol vessels in the naïve limbus and conjunctiva.Ocul Surf. 2020 Apr;18(2):277-285. doi: 10.1016/j.jtos.2020.02.005. Epub 2020 Feb 25. Ocul Surf. 2020. PMID: 32109562 Free PMC article.
-
Resistance to lethal ectromelia virus infection requires Type I interferon receptor in natural killer cells and monocytes but not in adaptive immune or parenchymal cells.PLoS Pathog. 2021 May 20;17(5):e1009593. doi: 10.1371/journal.ppat.1009593. eCollection 2021 May. PLoS Pathog. 2021. PMID: 34015056 Free PMC article.
-
The role of plasmacytoid dendritic cells (pDCs) in immunity during viral infections and beyond.Cell Mol Immunol. 2024 Sep;21(9):1008-1035. doi: 10.1038/s41423-024-01167-5. Epub 2024 May 22. Cell Mol Immunol. 2024. PMID: 38777879 Free PMC article. Review.
References
-
- Sei JJ, Haskett S, Kaminsky LW, Lin E, Truckenmiller ME, Bellone CJ, Buller RM, Norbury CC. 2015. Peptide-MHC-I from endogenous antigen outnumber those from exogenous antigen, irrespective of APC phenotype or activation. PLoS Pathog 11:e1004941. doi:10.1371/journal.ppat.1004941. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI070537/AI/NIAID NIH HHS/United States
- AI115230/AI/NIAID NIH HHS/United States
- R21 AI115230/AI/NIAID NIH HHS/United States
- R56 AI056094/AI/NIAID NIH HHS/United States
- T32 CA060395/CA/NCI NIH HHS/United States
- R01 AI056094/AI/NIAID NIH HHS/United States
- AI056094/AI/NIAID NIH HHS/United States
- AI083008/AI/NIAID NIH HHS/United States
- R21 AI097787/AI/NIAID NIH HHS/United States
- 5 T32CA60395-15/CA/NCI NIH HHS/United States
- AI097787/AI/NIAID NIH HHS/United States
- U19 AI083008/AI/NIAID NIH HHS/United States
- AI070537/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
