

Synthetic Long Peptide Derived from Mycobacterium tuberculosis Latency Antigen Rv1733c Protects against Tuberculosis
- PMID: 26202436
- PMCID: PMC4550671
- DOI: 10.1128/CVI.00271-15
Synthetic Long Peptide Derived from Mycobacterium tuberculosis Latency Antigen Rv1733c Protects against Tuberculosis
Abstract
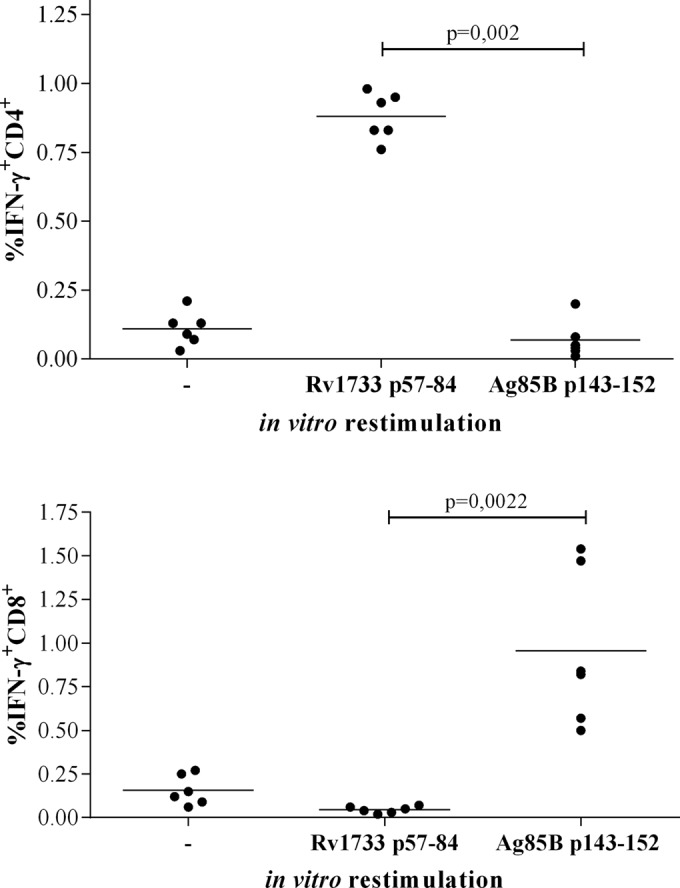
Responsible for 9 million new cases of active disease and nearly 2 million deaths each year, tuberculosis (TB) remains a global health threat of overwhelming dimensions. Mycobacterium bovis BCG, the only licensed vaccine available, fails to confer lifelong protection and to prevent reactivation of latent infection. Although 15 new vaccine candidates are now in clinical trials, an effective vaccine against TB remains elusive, and new strategies for vaccination are vital. BCG vaccination fails to induce immunity against Mycobacterium tuberculosis latency antigens. Synthetic long peptides (SLPs) combined with adjuvants have been studied mostly for therapeutic cancer vaccines, yet not for TB, and proved to induce efficient antitumor immunity. This study investigated an SLP derived from Rv1733c, a major M. tuberculosis latency antigen which is highly expressed by "dormant" M. tuberculosis and well recognized by T cells from latently M. tuberculosis-infected individuals. In order to assess its in vivo immunogenicity and protective capacity, Rv1733c SLP in CpG was administered to HLA-DR3 transgenic mice. Immunization with Rv1733c SLP elicited gamma interferon-positive/tumor necrosis factor-positive (IFN-γ(+)/TNF(+)) and IFN-γ(+) CD4(+) T cells and Rv1733c-specific antibodies and led to a significant reduction in the bacterial load in the lungs of M. tuberculosis-challenged mice. This was observed both in a pre- and in a post-M. tuberculosis challenge setting. Moreover, Rv1733c SLP immunization significantly boosted the protective efficacy of BCG, demonstrating the potential of M. tuberculosis latency antigens to improve BCG efficacy. These data suggest a promising role for M. tuberculosis latency antigen Rv1733c-derived SLPs as a novel TB vaccine approach, both in a prophylactic and in a postinfection setting.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures





Similar articles
-
A multistage-polyepitope vaccine protects against Mycobacterium tuberculosis infection in HLA-DR3 transgenic mice.Vaccine. 2012 Dec 14;30(52):7513-21. doi: 10.1016/j.vaccine.2012.10.045. Epub 2012 Oct 24. Vaccine. 2012. PMID: 23103299
-
Protective Vaccine Efficacy of the Complete Form of PPE39 Protein from Mycobacterium tuberculosis Beijing/K Strain in Mice.Clin Vaccine Immunol. 2017 Nov 6;24(11):e00219-17. doi: 10.1128/CVI.00219-17. Print 2017 Nov. Clin Vaccine Immunol. 2017. PMID: 28877927 Free PMC article.
-
Prime-boost vaccination strategy with bacillus Calmette-Guérin (BCG) and liposomized alpha-crystalline protein 1 reinvigorates BCG potency.Clin Exp Immunol. 2015 Aug;181(2):286-96. doi: 10.1111/cei.12634. Epub 2015 Jun 3. Clin Exp Immunol. 2015. PMID: 25845290 Free PMC article.
-
Immunogenic potential of latency associated antigens against Mycobacterium tuberculosis.Vaccine. 2014 Feb 3;32(6):712-6. doi: 10.1016/j.vaccine.2013.11.065. Epub 2013 Dec 2. Vaccine. 2014. PMID: 24300592 Review.
-
[Novel vaccines against M. tuberculosis].Kekkaku. 2006 Dec;81(12):745-51. Kekkaku. 2006. PMID: 17240920 Review. Japanese.
Cited by
-
Gene Regulatory Mechanism of Mycobacterium Tuberculosis during Dormancy.Curr Issues Mol Biol. 2024 Jun 11;46(6):5825-5844. doi: 10.3390/cimb46060348. Curr Issues Mol Biol. 2024. PMID: 38921019 Free PMC article. Review.
-
A Convenient Synthetic Method to Improve Immunogenicity of Mycobacterium tuberculosis Related T-Cell Epitope Peptides.Vaccines (Basel). 2019 Aug 27;7(3):101. doi: 10.3390/vaccines7030101. Vaccines (Basel). 2019. PMID: 31461944 Free PMC article.
-
Mycobacterium tuberculosis lineage 4 comprises globally distributed and geographically restricted sublineages.Nat Genet. 2016 Dec;48(12):1535-1543. doi: 10.1038/ng.3704. Epub 2016 Oct 31. Nat Genet. 2016. PMID: 27798628 Free PMC article.
-
Bridging the gaps to overcome major hurdles in the development of next-generation tuberculosis vaccines.Front Immunol. 2023 Aug 11;14:1193058. doi: 10.3389/fimmu.2023.1193058. eCollection 2023. Front Immunol. 2023. PMID: 37638056 Free PMC article. Review.
-
Photochemically-Mediated Inflammation and Cross-Presentation of Mycobacterium bovis BCG Proteins Stimulates Strong CD4 and CD8 T-Cell Responses in Mice.Front Immunol. 2022 Jan 31;13:815609. doi: 10.3389/fimmu.2022.815609. eCollection 2022. Front Immunol. 2022. PMID: 35173729 Free PMC article.
References
-
- World Health Organization. 2014. Global tuberculosis report 2014. World Health Organization, Geneva, Switzerland.
-
- Stop TB Partnership. 2011. The global plan to stop TB 2011–2015. Stop TB Partnership, Geneva, Switzerland.
-
- Roupie V, Romano M, Zhang L, Korf H, Lin MY, Franken KL, Ottenhoff TH, Klein MR, Huygen K. 2007. Immunogenicity of eight dormancy regulon-encoded proteins of Mycobacterium tuberculosis in DNA-vaccinated and tuberculosis-infected mice. Infect Immun 75:941–949. doi:10.1128/IAI.01137-06. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
