

SUMOylation at K707 of DGCR8 controls direct function of primary microRNA
- PMID: 26202964
- PMCID: PMC4652762
- DOI: 10.1093/nar/gkv741
SUMOylation at K707 of DGCR8 controls direct function of primary microRNA
Abstract
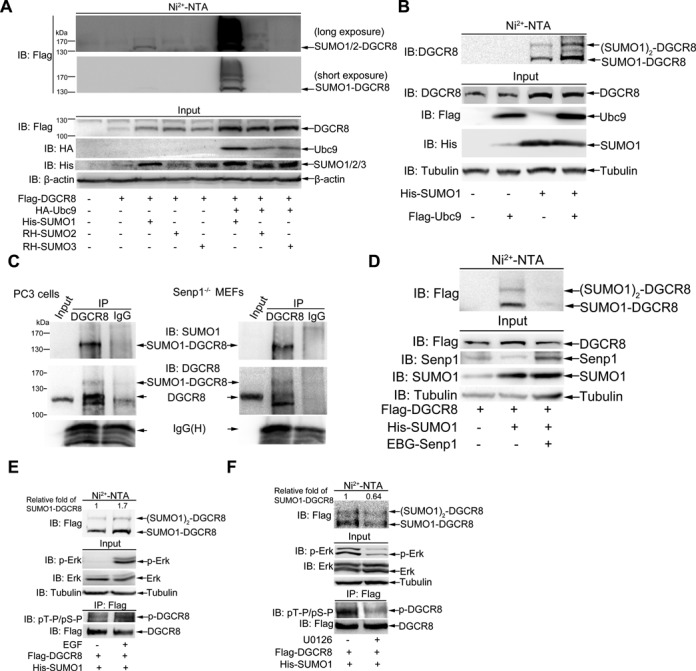
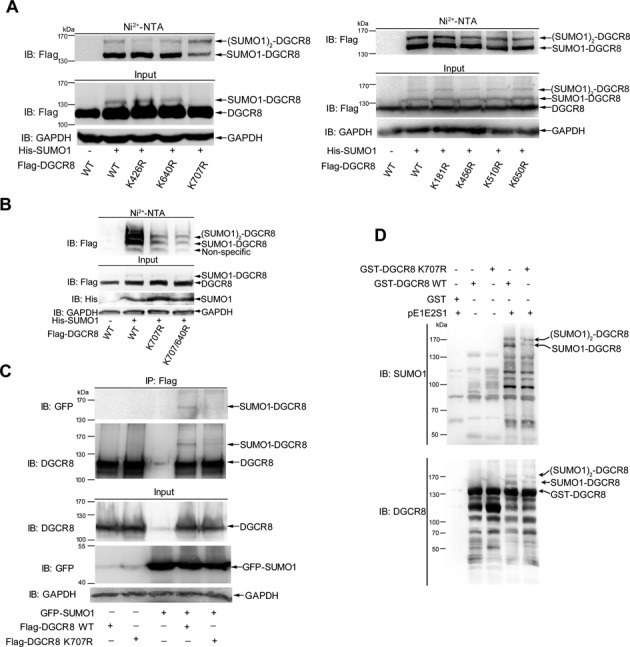
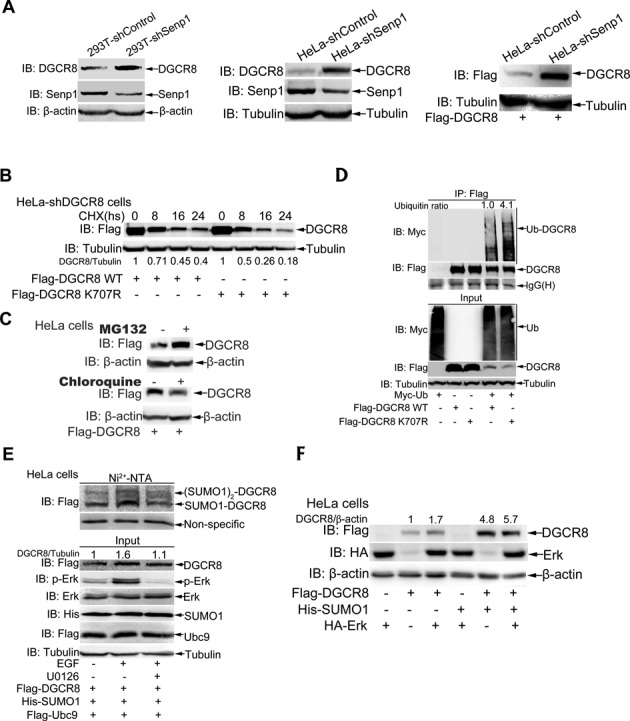
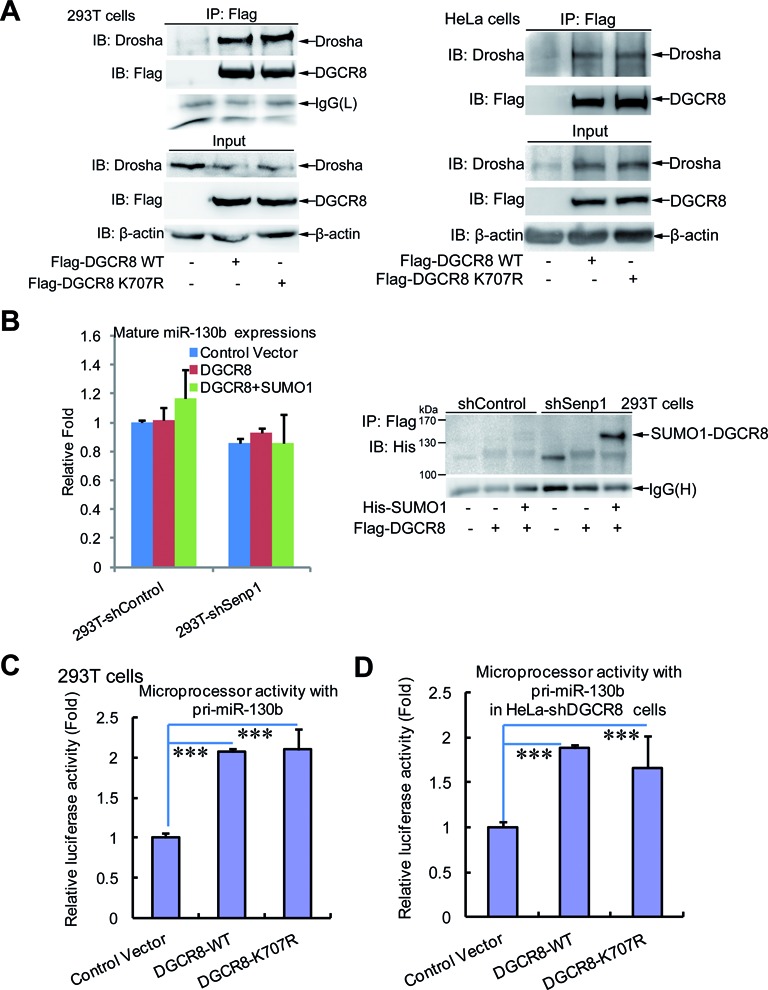
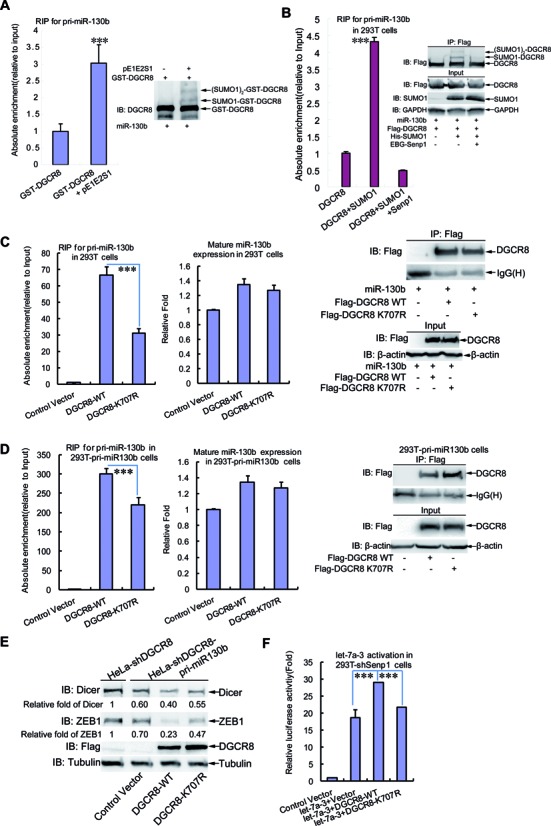
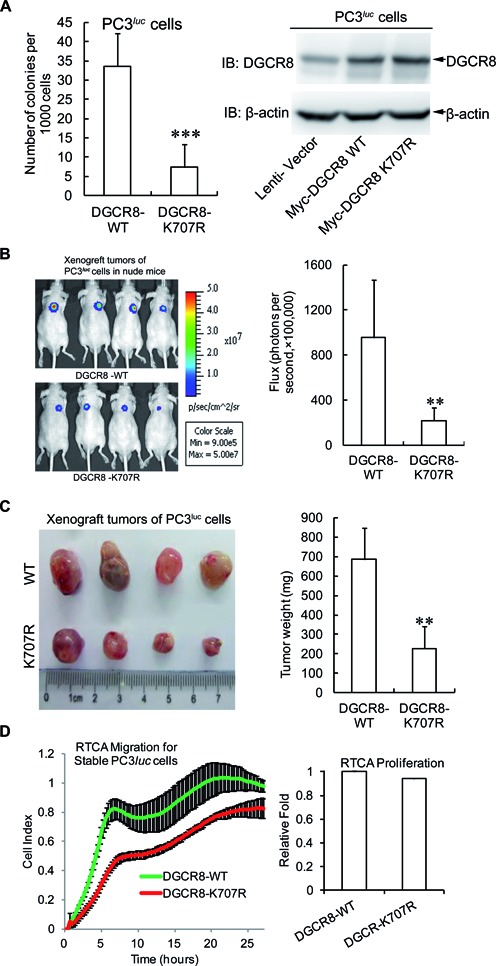
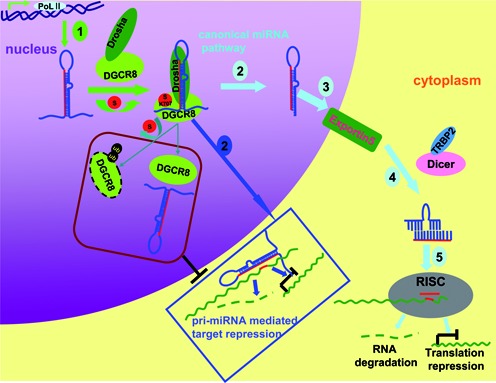
DGCR8 (DiGeorge syndrome critical region gene 8) is essential for primary microRNA (pri-miRNA) processing in the cell nucleus. It specifically combines with Drosha, a nuclear RNase III enzyme, to form the Microprocessor complex (MC) that cleaves pri-miRNA to precursor miRNA (pre-miRNA), which is further processed to mature miRNA by Dicer, a cytoplasmic RNase III enzyme. Increasing evidences suggest that pri-/pre-miRNAs have direct functions in regulation of gene expression, however the underlying mechanism how it is fine-tuned remains unclear. Here we find that DGCR8 is modified by SUMO1 at the major site K(707), which can be promoted by its ERK-activated phosphorylation. SUMOylation of DGCR8 enhances the protein stability by preventing the degradation via the ubiquitin proteasome pathway. More importantly, SUMOylation of DGCR8 does not alter its association with Drosha, the MC activity and miRNA biogenesis, but rather influences its affinity with pri-miRNAs. This altered affinity of DGCR8 with pri-miRNAs seems to control the direct functions of pri-miRNAs in recognition and repression of the target mRNAs, which is evidently linked to the DGCR8 function in regulation of tumorigenesis and cell migration. Collectively, our data suggest a novel mechanism that SUMOylation of DGCR8 controls direct functions of pri-miRNAs in gene silencing.
© The Author(s) 2015. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
SUMO1 modification of KHSRP regulates tumorigenesis by preventing the TL-G-Rich miRNA biogenesis.Mol Cancer. 2017 Oct 11;16(1):157. doi: 10.1186/s12943-017-0724-6. Mol Cancer. 2017. PMID: 29020972 Free PMC article.
-
The Ubiquitin-specific Protease USP36 Associates with the Microprocessor Complex and Regulates miRNA Biogenesis by SUMOylating DGCR8.Cancer Res Commun. 2023 Mar 20;3(3):459-470. doi: 10.1158/2767-9764.CRC-22-0344. eCollection 2023 Mar. Cancer Res Commun. 2023. PMID: 36950067 Free PMC article.
-
Characterization of DGCR8/Pasha, the essential cofactor for Drosha in primary miRNA processing.Nucleic Acids Res. 2006;34(16):4622-9. doi: 10.1093/nar/gkl458. Epub 2006 Sep 8. Nucleic Acids Res. 2006. PMID: 16963499 Free PMC article.
-
MicroRNA biogenesis: isolation and characterization of the microprocessor complex.Methods Mol Biol. 2006;342:33-47. doi: 10.1385/1-59745-123-1:33. Methods Mol Biol. 2006. PMID: 16957365 Review.
-
Autoregulatory mechanisms controlling the Microprocessor.Adv Exp Med Biol. 2010;700:56-66. Adv Exp Med Biol. 2010. PMID: 21627030 Review.
Cited by
-
SUMOylation of the m6A-RNA methyltransferase METTL3 modulates its function.Nucleic Acids Res. 2018 Jun 1;46(10):5195-5208. doi: 10.1093/nar/gky156. Nucleic Acids Res. 2018. PMID: 29506078 Free PMC article.
-
Regulation of MicroRNAs.Methods Mol Biol. 2022;2257:1-32. doi: 10.1007/978-1-0716-1170-8_1. Methods Mol Biol. 2022. PMID: 34432271
-
STAT5A induced LINC01198 promotes proliferation of glioma cells through stabilizing DGCR8.Aging (Albany NY). 2020 Apr 4;12(7):5675-5692. doi: 10.18632/aging.102938. Epub 2020 Apr 4. Aging (Albany NY). 2020. PMID: 32246817 Free PMC article.
-
SUMOylation modulates the LIN28A-let-7 signaling pathway in response to cellular stresses in cancer cells.Mol Oncol. 2020 Sep;14(9):2288-2312. doi: 10.1002/1878-0261.12694. Epub 2020 Jun 1. Mol Oncol. 2020. PMID: 32333719 Free PMC article.
-
Imbalance of a KLF4-miR-7 auto-regulatory feedback loop promotes prostate cancer cell growth by impairing microRNA processing.Am J Cancer Res. 2018 Feb 1;8(2):226-244. eCollection 2018. Am J Cancer Res. 2018. PMID: 29511594 Free PMC article.
References
-
- Landthaler M., Yalcin A., Tuschl T. The human DiGeorge syndrome critical region gene 8 and Its D. melanogaster homolog are required for miRNA biogenesis. Curr. Biol. 2004;14:2162–2167. - PubMed
-
- Tomari Y., Zamore P.D. MicroRNA biogenesis: drosha can't cut it without a partner. Curr. Biol. 2005;15:R61–R64. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
