

Regulation of ERK-MAPK signaling in human epidermis
- PMID: 26209520
- PMCID: PMC4514964
- DOI: 10.1186/s12918-015-0187-6
Regulation of ERK-MAPK signaling in human epidermis
Abstract
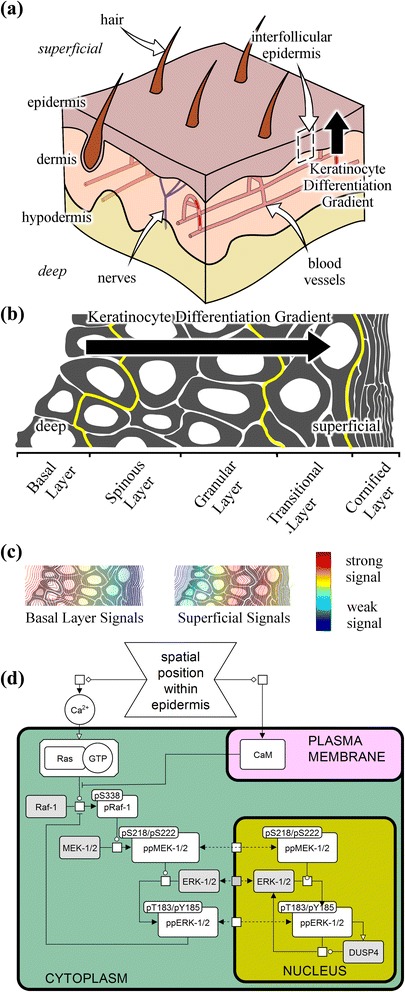
Background: The skin is largely comprised of keratinocytes within the interfollicular epidermis. Over approximately two weeks these cells differentiate and traverse the thickness of the skin. The stage of differentiation is therefore reflected in the positions of cells within the tissue, providing a convenient axis along which to study the signaling events that occur in situ during keratinocyte terminal differentiation, over this extended two-week timescale. The canonical ERK-MAPK signaling cascade (Raf-1, MEK-1/2 and ERK-1/2) has been implicated in controlling diverse cellular behaviors, including proliferation and differentiation. While the molecular interactions involved in signal transduction through this cascade have been well characterized in cell culture experiments, our understanding of how this sequence of events unfolds to determine cell fate within a homeostatic tissue environment has not been fully characterized.
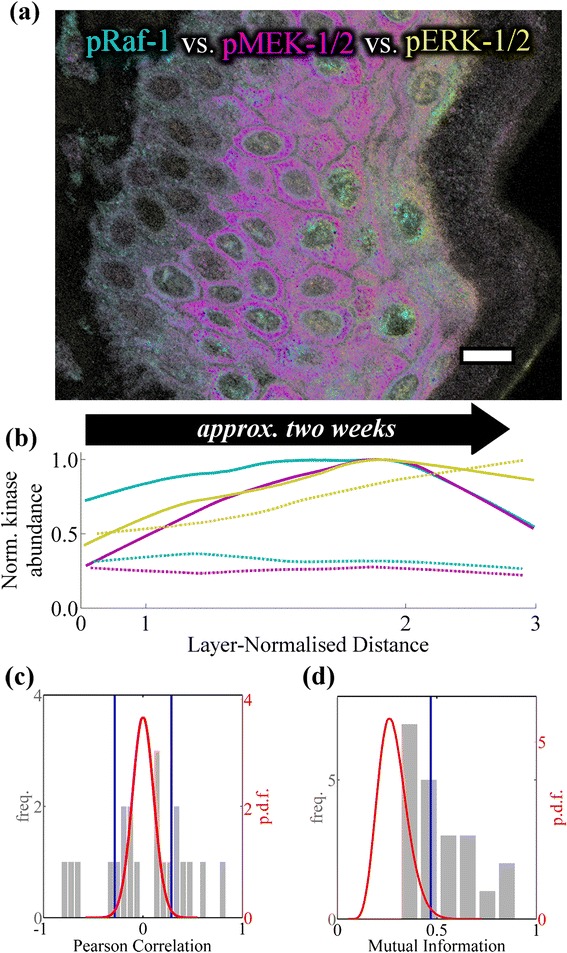
Methods: We measured the abundance of total and phosphorylated ERK-MAPK signaling proteins within interfollicular keratinocytes in transverse cross-sections of human epidermis using immunofluorescence microscopy. To investigate these data we developed a mathematical model of the signaling cascade using a normalized-Hill differential equation formalism.
Results: These data show coordinated variation in the abundance of phosphorylated ERK-MAPK components across the epidermis. Statistical analysis of these data shows that associations between phosphorylated ERK-MAPK components which correspond to canonical molecular interactions are dependent upon spatial position within the epidermis. The model demonstrates that the spatial profile of activation for ERK-MAPK signaling components across the epidermis may be maintained in a cell-autonomous fashion by an underlying spatial gradient in calcium signaling.
Conclusions: Our data demonstrate an extended phospho-protein profile of ERK-MAPK signaling cascade components across the epidermis in situ, and statistical associations in these data indicate canonical ERK-MAPK interactions underlie this spatial profile of ERK-MAPK activation. Using mathematical modelling we have demonstrated that spatially varying calcium signaling components across the epidermis may be sufficient to maintain the spatial profile of ERK-MAPK signaling cascade components in a cell-autonomous manner. These findings may have significant implications for the wide range of cancer drugs which therapeutically target ERK-MAPK signaling components.
Figures


References
-
- Eckert RL, Crish JF, Robinson NA. The epidermal keratinocyte as a model for the study of gene regulation and cell differentiation. Physiol Rev. 1997;77:397–424. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
