

Fanconi Anemia Mesenchymal Stromal Cells-Derived Glycerophospholipids Skew Hematopoietic Stem Cell Differentiation Through Toll-Like Receptor Signaling
- PMID: 26212365
- PMCID: PMC4618082
- DOI: 10.1002/stem.2100
Fanconi Anemia Mesenchymal Stromal Cells-Derived Glycerophospholipids Skew Hematopoietic Stem Cell Differentiation Through Toll-Like Receptor Signaling
Abstract
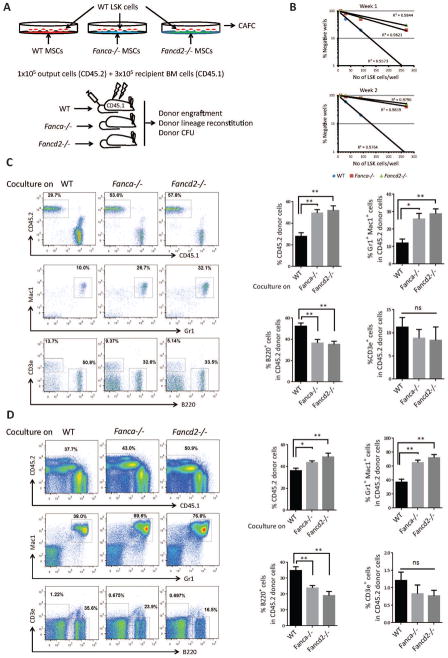
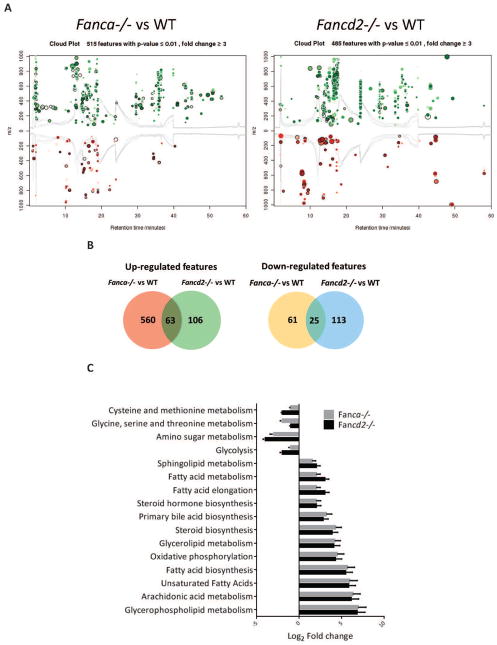
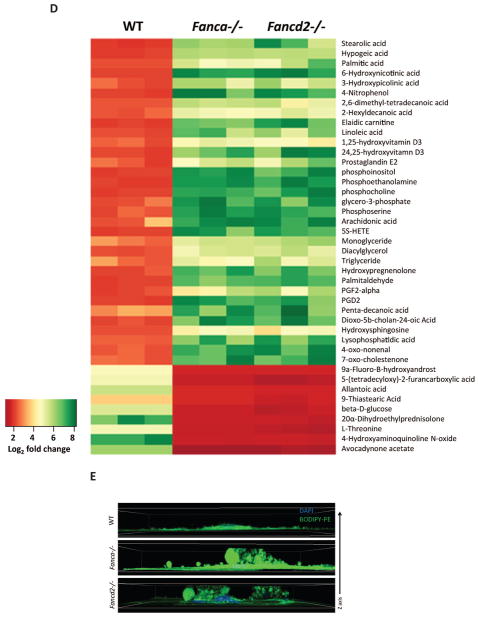
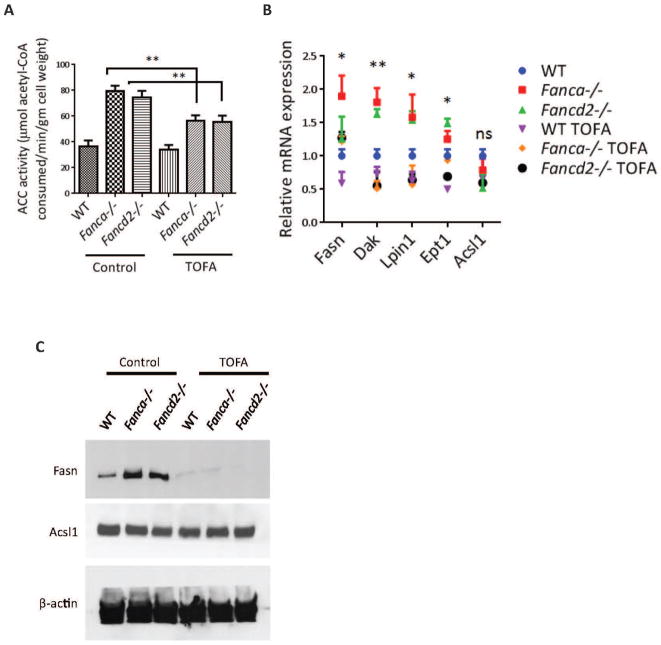
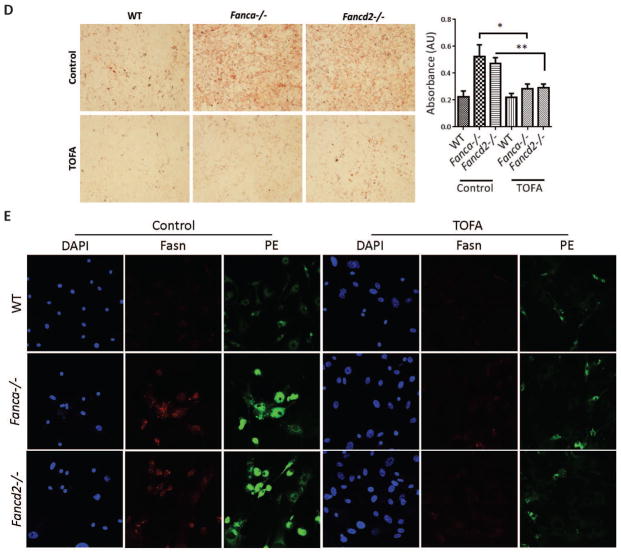
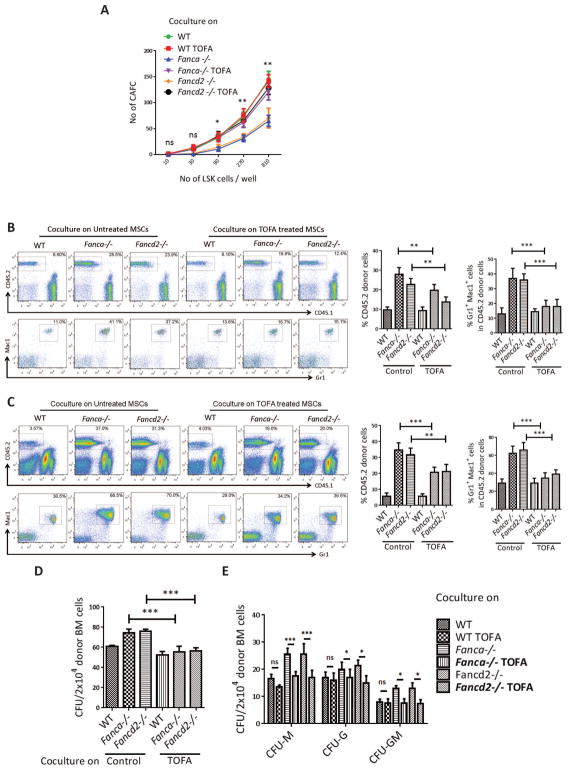
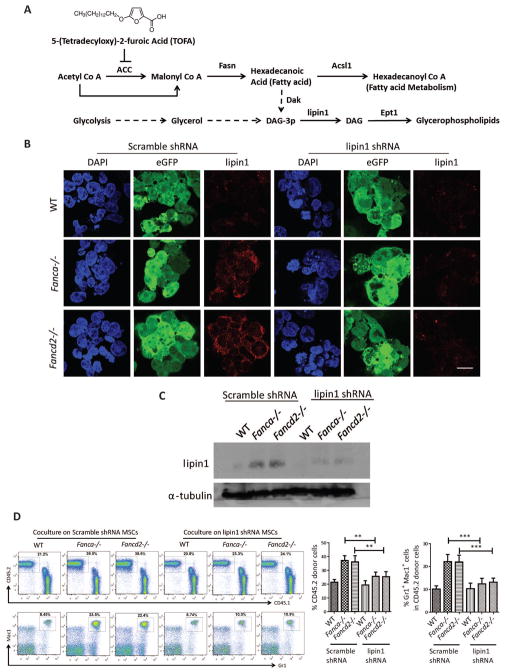
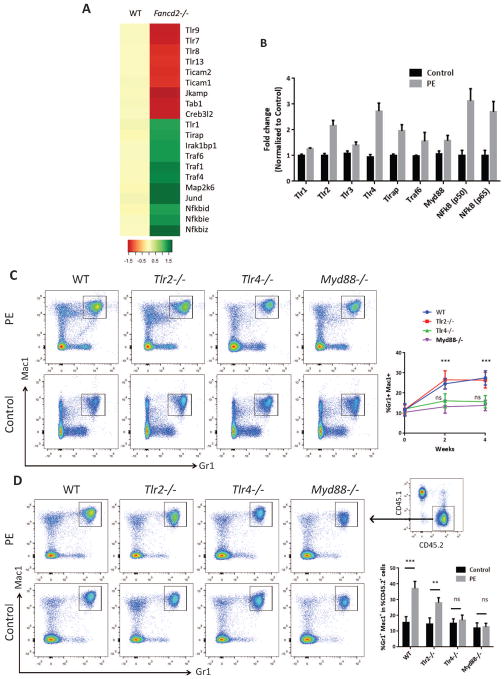
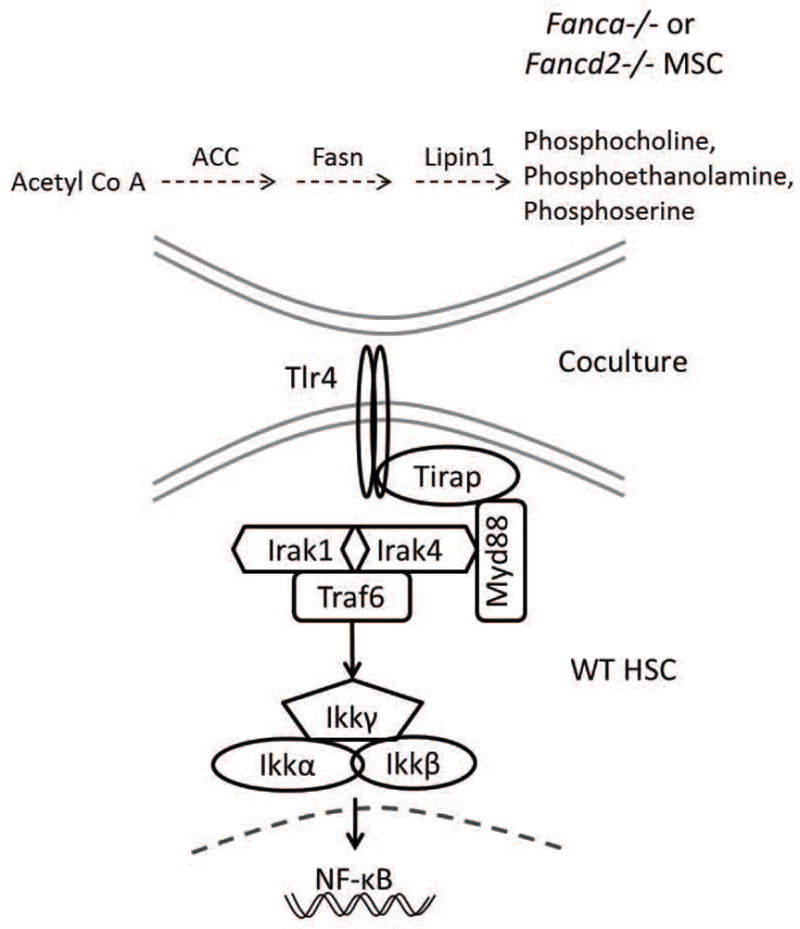
Fanconi anemia (FA) patients develop bone marrow (BM) failure or leukemia. One standard care for these devastating complications is hematopoietic stem cell transplantation. We identified a group of mesenchymal stromal cells (MSCs)-derived metabolites, glycerophospholipids, and their endogenous inhibitor, 5-(tetradecyloxy)-2-furoic acid (TOFA), as regulators of donor hematopoietic stem and progenitor cells. We provided two pieces of evidence that TOFA could improve hematopoiesis-supporting function of FA MSCs: (a) limiting-dilution cobblestone area-forming cell assay revealed that TOFA significantly increased cobblestone colonies in Fanca-/- or Fancd2-/- cocultures compared to untreated cocultures. (b) Competitive repopulating assay using output cells collected from cocultures showed that TOFA greatly alleviated the abnormal expansion of the donor myeloid (CD45.2+Gr1+Mac1+) compartment in both peripheral blood and BM of recipient mice transplanted with cells from Fanca-/- or Fancd2-/- cocultures. Furthermore, mechanistic studies identified Tlr4 signaling as the responsible pathway mediating the effect of glycerophospholipids. Thus, targeting glycerophospholipid biosynthesis in FA MSCs could be a therapeutic strategy to improve hematopoiesis and stem cell transplantation.
Keywords: Fanconi anemia; Hematopoietic stem cell transplantation; Marrow stromal stem cells; Myeloid cells.
© 2015 AlphaMed Press.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures
References
-
- Bagby GC., Jr Genetic basis of Fanconi anemia. Curr Opin Hematol. 2003;10(1):68–76. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
