

Characterization of RNA silencing components in the plant pathogenic fungus Fusarium graminearum
- PMID: 26212591
- PMCID: PMC4515635
- DOI: 10.1038/srep12500
Characterization of RNA silencing components in the plant pathogenic fungus Fusarium graminearum
Abstract
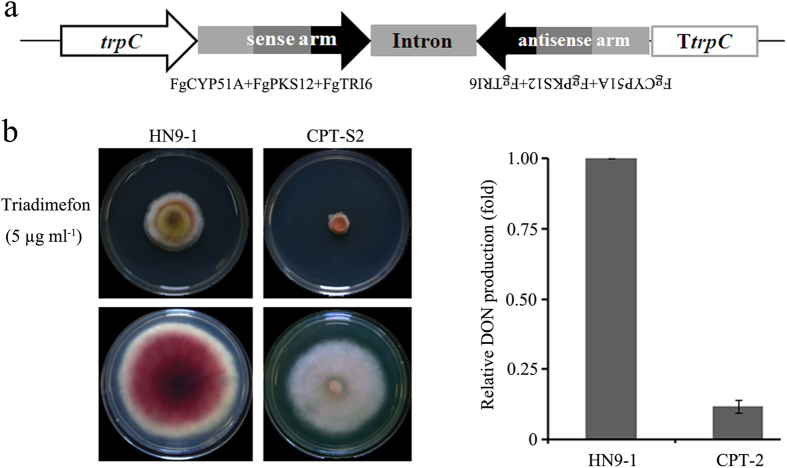
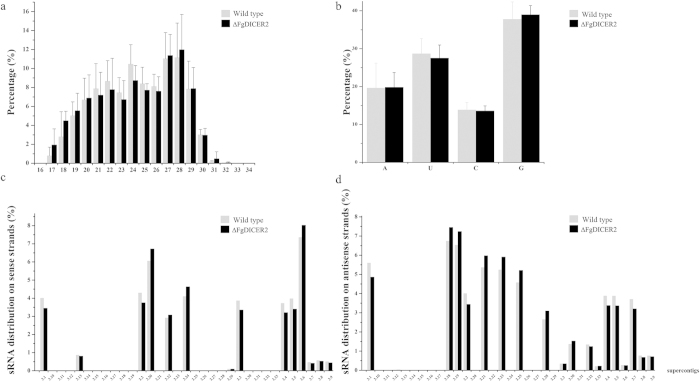
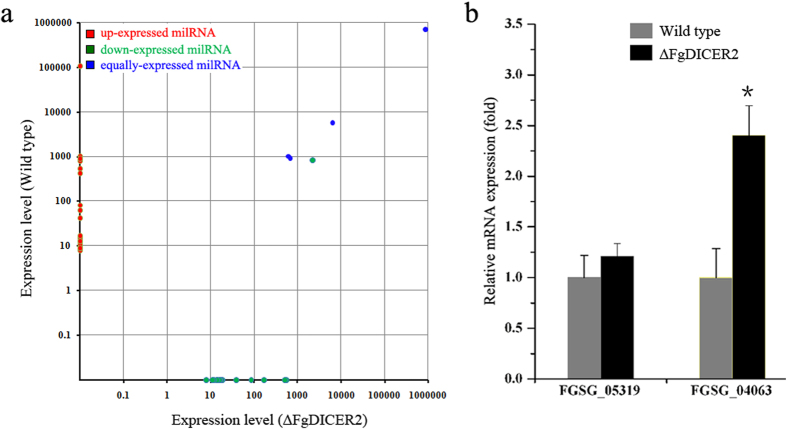
The RNA interference (RNAi) plays a critical role in gene regulation in a variety of eukaryotic organisms. However, the role of RNAi remains largely unclear in plant pathogenic fungi. In this study, we explored the roles of core components of the RNAi pathway in Fusarium graminearum, the major causal agent of wheat head blight. Our results demonstrated that the hairpin RNA (hpRNA) can efficiently silence the expression level of target gene, and the argonaute protein FgAgo1 and dicer protein FgDicer2 are important in this silencing process. RNAi machinery was not involved in growth, abiotic stress and pathogenesis in F. graminearum under tested conditions. We firstly applied high-throughput sequencing technology to elucidate small RNA (17-40 nucleotides) (sRNA) transcriptome in F. graminearum, and found that a total of forty-nine micro-like-RNA (milRNA) candidates were identified in the wild-type and ∆FgDICER2, and twenty-four of them were FgDicer2-dependent. Fg-milRNA-4 negatively regulated expression of its target gene. Taken together, our results indicated that the hpRNA-induced gene silencing was a valuable genetic tool for exploring gene function in F. graminearum. FgAgo1 and FgDicer2 proteins played a critical role in the hpRNA mediated gene silencing process. In addition, FgDicer2 was involved in sRNA transcription and milRNA generation in this fungus.
Figures



References
-
- Carreras-Villasenor N., Esquivel-Naranjo E. U., Villalobos-Escobedo J. M., Abreu-Goodger C. & Herrera-Estrella A. The RNAi machinery regulates growth and development in the filamentous fungus Trichoderma atroviride. Mol. Microbiol. 89, 96–112 (2013). - PubMed
-
- Brodersen P. & Voinnet O. The diversity of RNA silencing pathways in plants. Trends Genet. 22, 268–280 (2006). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
