

Distinct Neural Circuits Control Rhythm Inhibition and Spitting by the Myogenic Pharynx of C. elegans
- PMID: 26212880
- PMCID: PMC4546535
- DOI: 10.1016/j.cub.2015.06.052
Distinct Neural Circuits Control Rhythm Inhibition and Spitting by the Myogenic Pharynx of C. elegans
Abstract
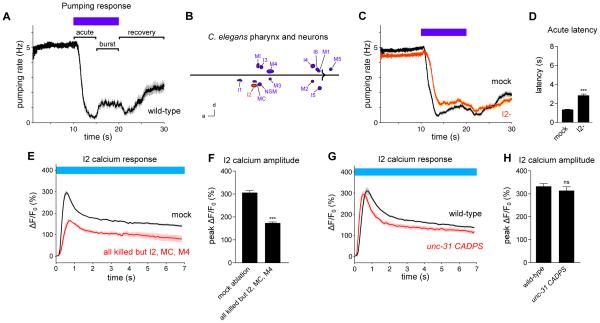
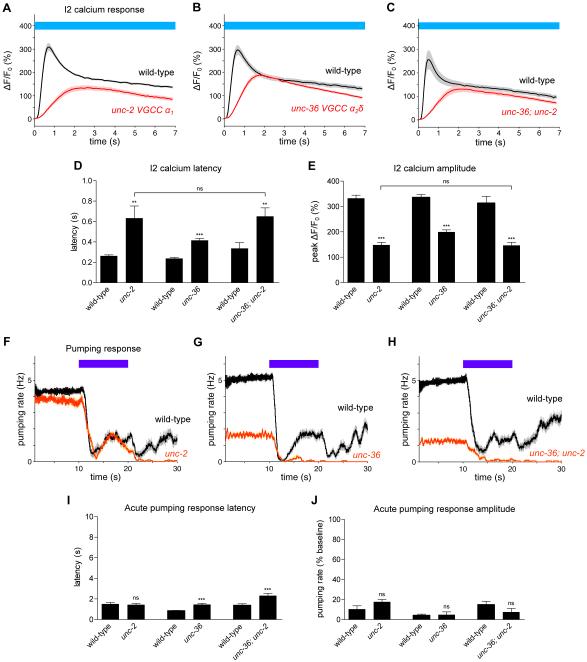
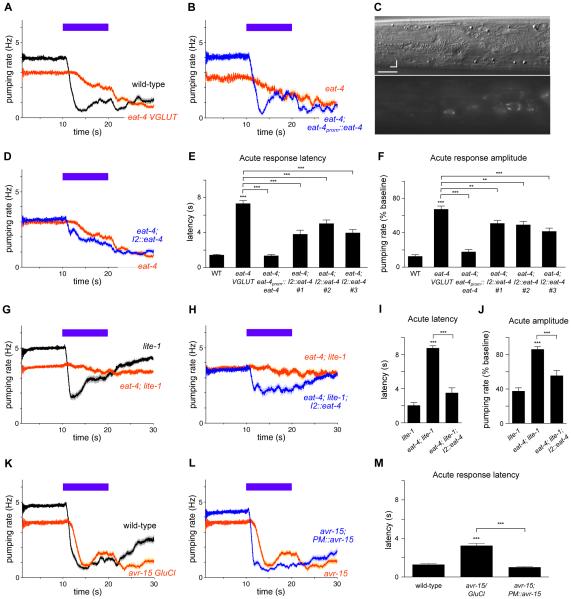
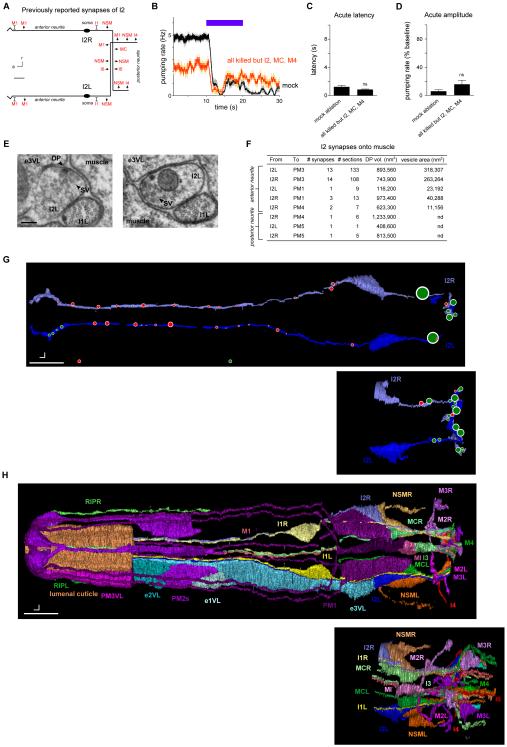
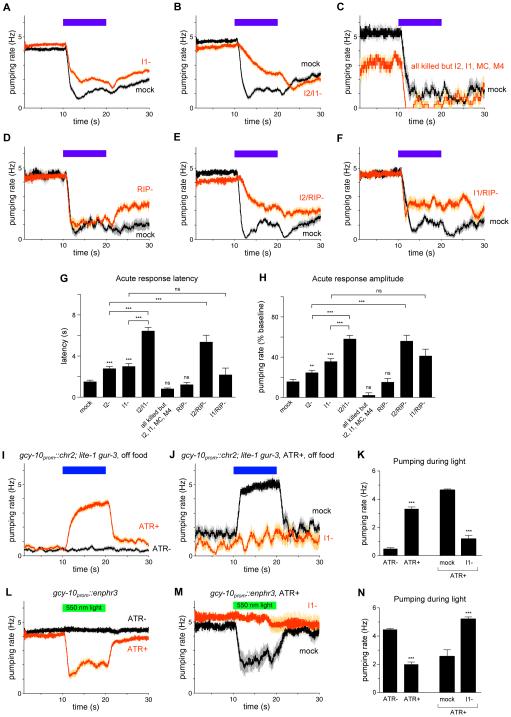
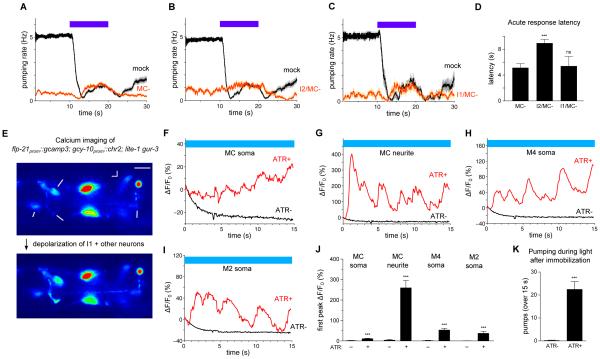
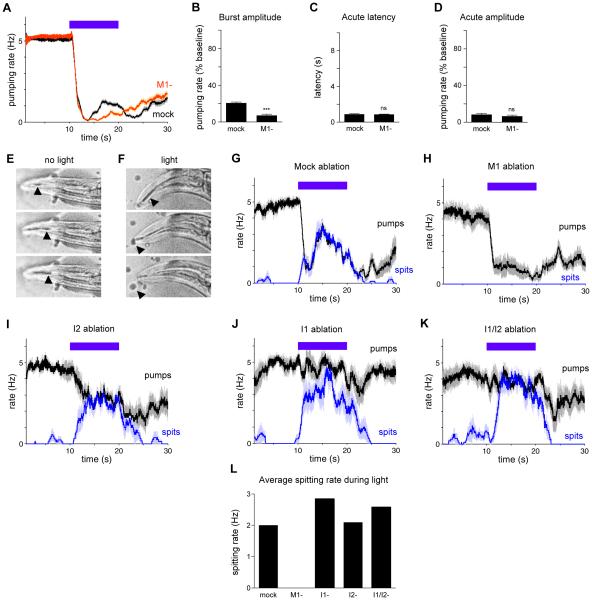
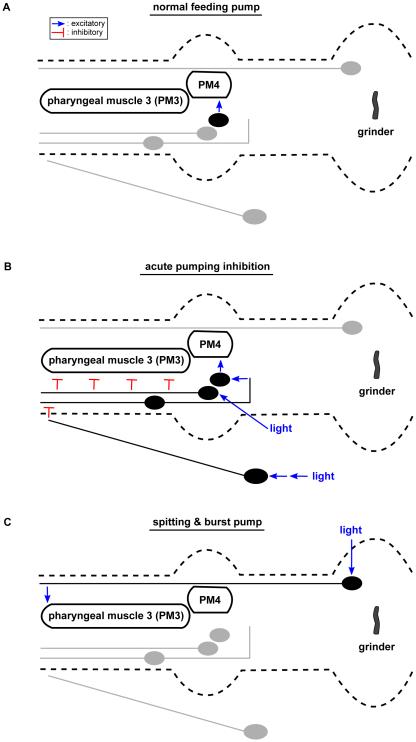
Neural circuits have long been known to modulate myogenic muscles such as the heart, yet a mechanistic understanding at the cellular and molecular levels remains limited. We studied how light inhibits pumping of the Caenorhabditis elegans pharynx, a myogenic muscular pump for feeding, and found three neural circuits that alter pumping. First, light inhibits pumping via the I2 neuron monosynaptic circuit. Our electron microscopic reconstruction of the anterior pharynx revealed evidence for synapses from I2 onto muscle that were missing from the published connectome, and we show that these "missed synapses" are likely functional. Second, light inhibits pumping through the RIP-I1-MC neuron polysynaptic circuit, in which an inhibitory signal is likely transmitted from outside the pharynx into the pharynx in a manner analogous to how the mammalian autonomic nervous system controls the heart. Third, light causes a novel pharyngeal behavior, reversal of flow or "spitting," which is induced by the M1 neuron. These three neural circuits show that neurons can control a myogenic muscle organ not only by changing the contraction rate but also by altering the functional consequences of the contraction itself, transforming swallowing into spitting. Our observations also illustrate why connectome builders and users should be cognizant that functional synaptic connections might exist despite the absence of a declared synapse in the connectome.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
Comment in
-
Neural Circuits: From Structure to Function and Back.Curr Biol. 2015 Aug 17;25(16):R711-3. doi: 10.1016/j.cub.2015.06.048. Curr Biol. 2015. PMID: 26294182
References
-
- Irisawa H, Brown HF, Giles W. Cardiac pacemaking in the sinoatrial node. Physiological Reviews. 1993;73:197–227. - PubMed
-
- Donnelly G, Jackson TD, Ambrous K, Ye J, Safdar A, Farraway L, Huizinga JD. The myogenic component in distention-induced peristalsis in the guinea pig small intestine. American Journal of Physiology - Gastrointestinal and Liver Physiology. 2001;280:G491–G500. - PubMed
-
- Palma J-A, Benarroch EE. Neural control of the heart. Neurology. 2014;83:1–11. - PubMed
-
- Chalasani SH, Chronis N, Tsunozaki M, Gray JM, Ramot D, Goodman MB, Bargmann CI. Dissecting a circuit for olfactory behavior in Caenorhabditis elegans. Nature. 2007;450:63–70. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
