

Determinants of different deep and superficial CA1 pyramidal cell dynamics during sharp-wave ripples
- PMID: 26214372
- PMCID: PMC4820637
- DOI: 10.1038/nn.4074
Determinants of different deep and superficial CA1 pyramidal cell dynamics during sharp-wave ripples
Abstract
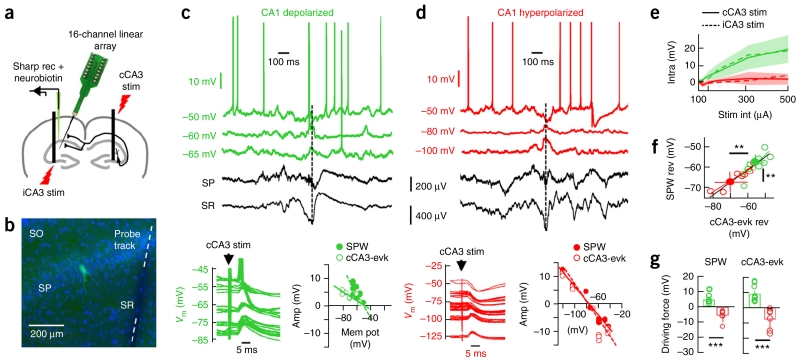
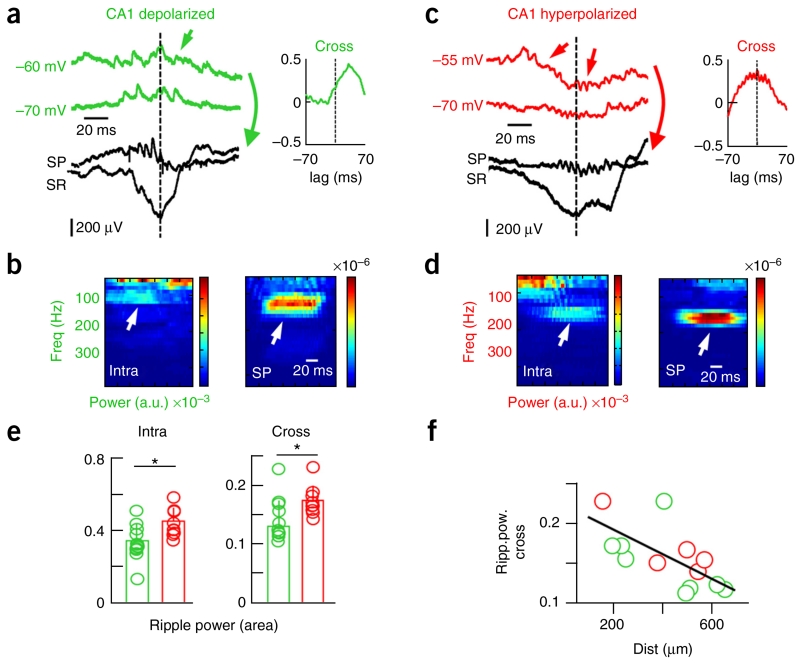
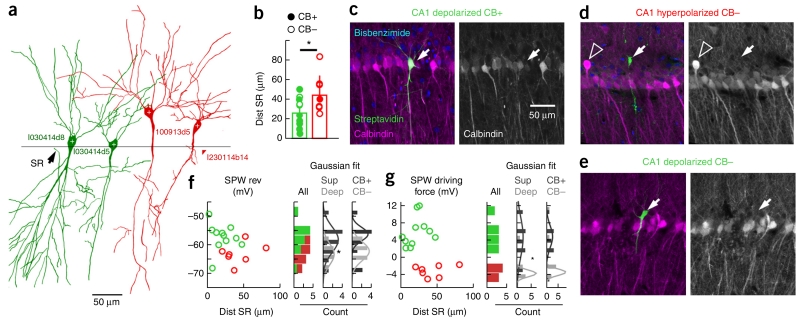
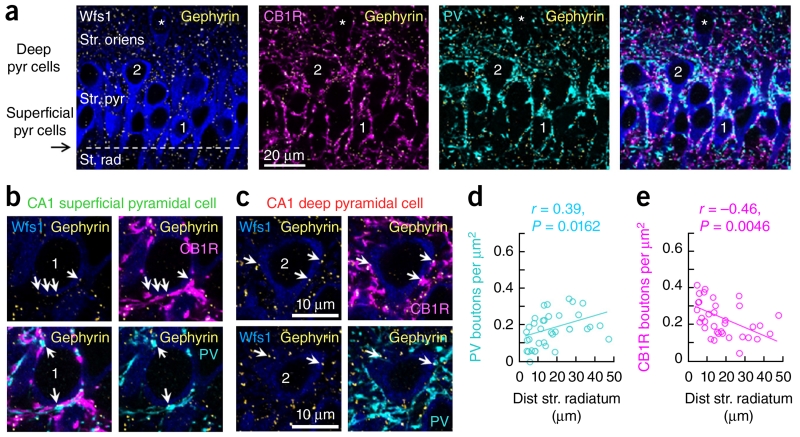
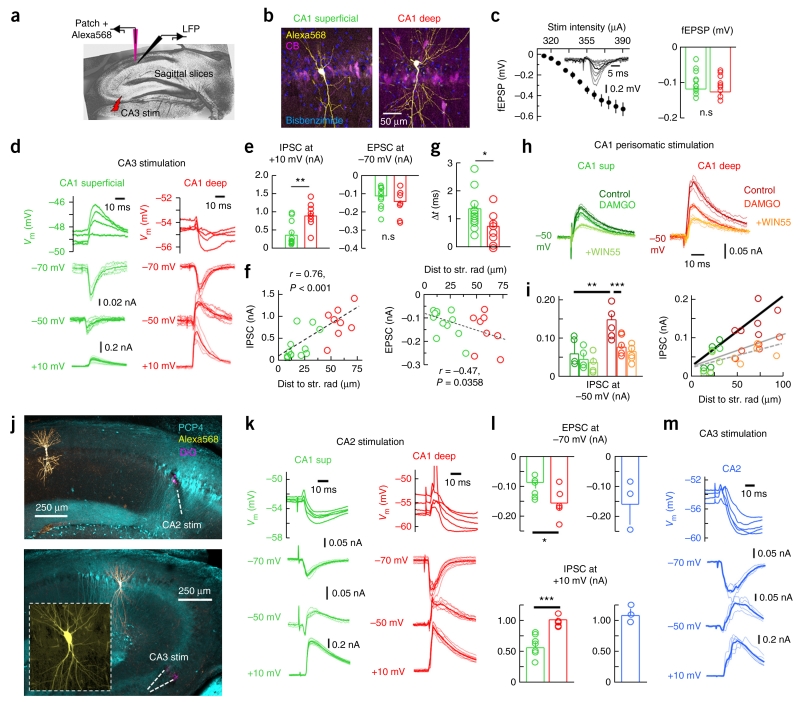
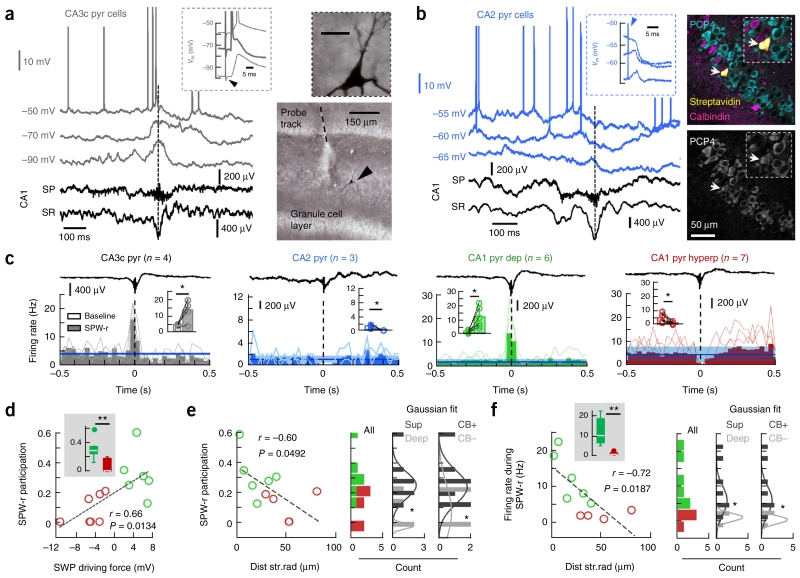
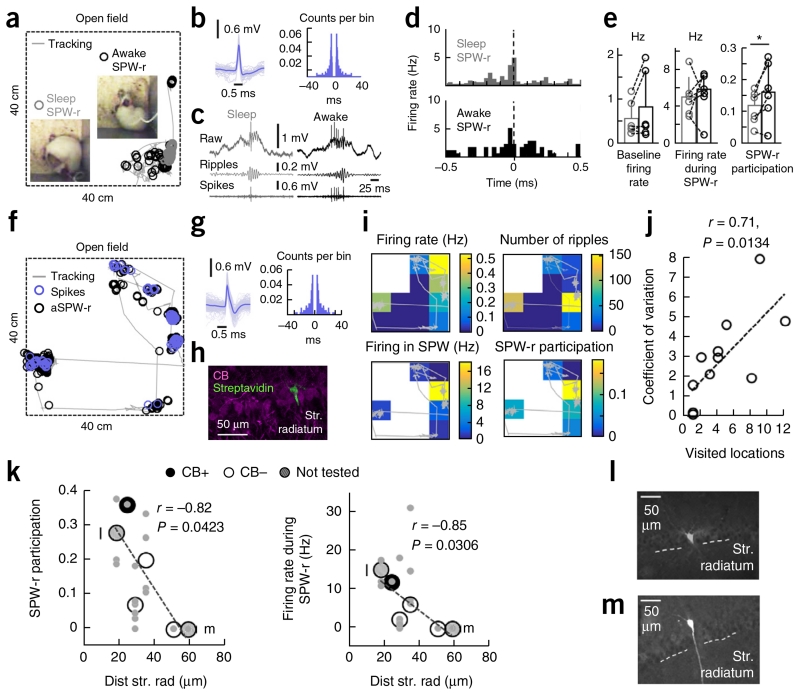
Sharp-wave ripples represent a prominent synchronous activity pattern in the mammalian hippocampus during sleep and immobility. GABAergic interneuronal types are silenced or fire during these events, but the mechanism of pyramidal cell (PC) participation remains elusive. We found opposite membrane polarization of deep (closer to stratum oriens) and superficial (closer to stratum radiatum) rat CA1 PCs during sharp-wave ripples. Using sharp and multi-site recordings in combination with neurochemical profiling, we observed a predominant inhibitory drive of deep calbindin (CB)-immunonegative PCs that contrasts with a prominent depolarization of superficial CB-immunopositive PCs. Biased contribution of perisomatic GABAergic inputs, together with suppression of CA2 PCs, may explain the selection of CA1 PCs during sharp-wave ripples. A deep-superficial gradient interacted with behavioral and spatial effects to determine cell participation during sleep and awake sharp-wave ripples in freely moving rats. Thus, the firing dynamics of hippocampal PCs are exquisitely controlled at subcellular and microcircuit levels in a cell type-selective manner.
Figures
References
-
- Buzsáki G, Leung LWS, Vanderwolf CH. Cellular bases of hippocampal EEG in the behaving rat. Brain Res. 1983;287:139–171. - PubMed
-
- Skaggs WE, et al. EEG sharp waves and sparse ensemble unit activity in the macaque hippocampus. J. Neurophysiol. 2007;98:898–910. - PubMed
-
- Csicsvari J, Hirase H, Mamiya A, Buzsáki G. Ensemble patterns of hippocampal CA3-CA1 neurons during sharp wave-associated population events. Neuron. 2000;28:585–594. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
