

GATA2 is required for lymphatic vessel valve development and maintenance
- PMID: 26214525
- PMCID: PMC4563742
- DOI: 10.1172/JCI78888
GATA2 is required for lymphatic vessel valve development and maintenance
Abstract
Heterozygous germline mutations in the zinc finger transcription factor GATA2 have recently been shown to underlie a range of clinical phenotypes, including Emberger syndrome, a disorder characterized by lymphedema and predisposition to myelodysplastic syndrome/acute myeloid leukemia (MDS/AML). Despite well-defined roles in hematopoiesis, the functions of GATA2 in the lymphatic vasculature and the mechanisms by which GATA2 mutations result in lymphedema have not been characterized. Here, we have provided a molecular explanation for lymphedema predisposition in a subset of patients with germline GATA2 mutations. Specifically, we demonstrated that Emberger-associated GATA2 missense mutations result in complete loss of GATA2 function, with respect to the capacity to regulate the transcription of genes that are important for lymphatic vessel valve development. We identified a putative enhancer element upstream of the key lymphatic transcriptional regulator PROX1 that is bound by GATA2, and the transcription factors FOXC2 and NFATC1. Emberger GATA2 missense mutants had a profoundly reduced capacity to bind this element. Conditional Gata2 deletion in mice revealed that GATA2 is required for both development and maintenance of lymphovenous and lymphatic vessel valves. Together, our data unveil essential roles for GATA2 in the lymphatic vasculature and explain why a select catalogue of human GATA2 mutations results in lymphedema.
Figures






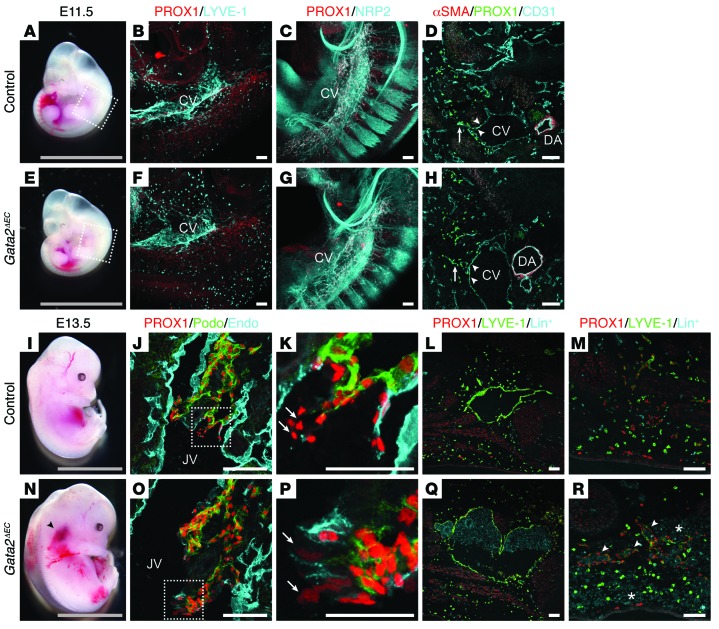

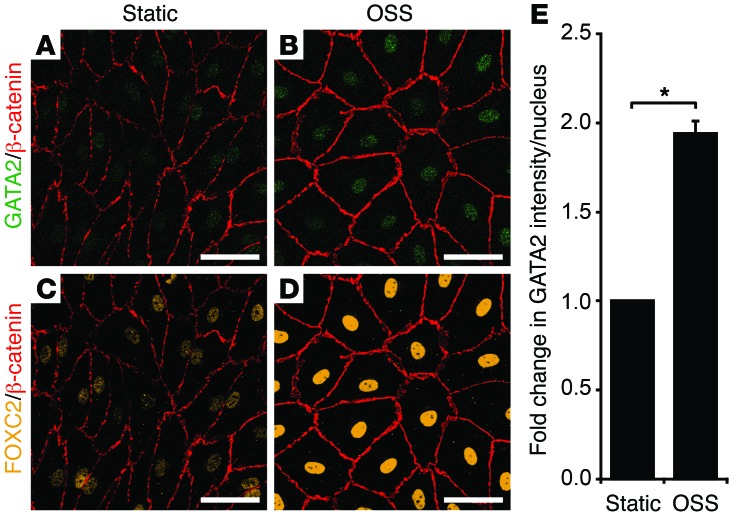




Comment in
-
Lymphatic vessel development: fluid flow and valve-forming cells.J Clin Invest. 2015 Aug 3;125(8):2924-6. doi: 10.1172/JCI83189. Epub 2015 Jul 27. J Clin Invest. 2015. PMID: 26214518 Free PMC article.
Similar articles
-
Loss-of-function germline GATA2 mutations in patients with MDS/AML or MonoMAC syndrome and primary lymphedema reveal a key role for GATA2 in the lymphatic vasculature.Blood. 2012 Feb 2;119(5):1283-91. doi: 10.1182/blood-2011-08-374363. Epub 2011 Dec 6. Blood. 2012. PMID: 22147895 Free PMC article.
-
Lymphatic vessel development: fluid flow and valve-forming cells.J Clin Invest. 2015 Aug 3;125(8):2924-6. doi: 10.1172/JCI83189. Epub 2015 Jul 27. J Clin Invest. 2015. PMID: 26214518 Free PMC article.
-
Complementary Wnt Sources Regulate Lymphatic Vascular Development via PROX1-Dependent Wnt/β-Catenin Signaling.Cell Rep. 2018 Oct 16;25(3):571-584.e5. doi: 10.1016/j.celrep.2018.09.049. Cell Rep. 2018. PMID: 30332639 Free PMC article.
-
[GATA2 deficiency].Nihon Rinsho. 2014 Oct;72(10):1861-9. Nihon Rinsho. 2014. PMID: 25509816 Review. Japanese.
-
Heterogeneity of GATA2-related myeloid neoplasms.Int J Hematol. 2017 Aug;106(2):175-182. doi: 10.1007/s12185-017-2285-2. Epub 2017 Jun 22. Int J Hematol. 2017. PMID: 28643018 Review.
Cited by
-
GATA2 regulates blood/lymph separation in a platelet-dependent and lymphovenous valve-independent manner.Microcirculation. 2023 Apr;30(2-3):e12787. doi: 10.1111/micc.12787. Epub 2022 Oct 19. Microcirculation. 2023. PMID: 36197446 Free PMC article.
-
GATA2 controls lymphatic endothelial cell junctional integrity and lymphovenous valve morphogenesis through miR-126.Development. 2019 Nov 5;146(21):dev184218. doi: 10.1242/dev.184218. Development. 2019. PMID: 31582413 Free PMC article.
-
RASA1 regulates the function of lymphatic vessel valves in mice.J Clin Invest. 2017 Jun 30;127(7):2569-2585. doi: 10.1172/JCI89607. Epub 2017 May 22. J Clin Invest. 2017. PMID: 28530642 Free PMC article.
-
Generation and application of endogenously floxed alleles for cell-specific knockout in zebrafish.Dev Cell. 2023 Nov 20;58(22):2614-2626.e7. doi: 10.1016/j.devcel.2023.07.022. Epub 2023 Aug 25. Dev Cell. 2023. PMID: 37633272 Free PMC article.
-
Fetal nuchal edema and developmental anomalies caused by gene mutations in mice.Front Cell Dev Biol. 2022 Aug 30;10:949013. doi: 10.3389/fcell.2022.949013. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 36111337 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
