

Alpha-, Delta- and PP-cells: Are They the Architectural Cornerstones of Islet Structure and Co-ordination?
- PMID: 26216135
- PMCID: PMC4530398
- DOI: 10.1369/0022155415583535
Alpha-, Delta- and PP-cells: Are They the Architectural Cornerstones of Islet Structure and Co-ordination?
Abstract
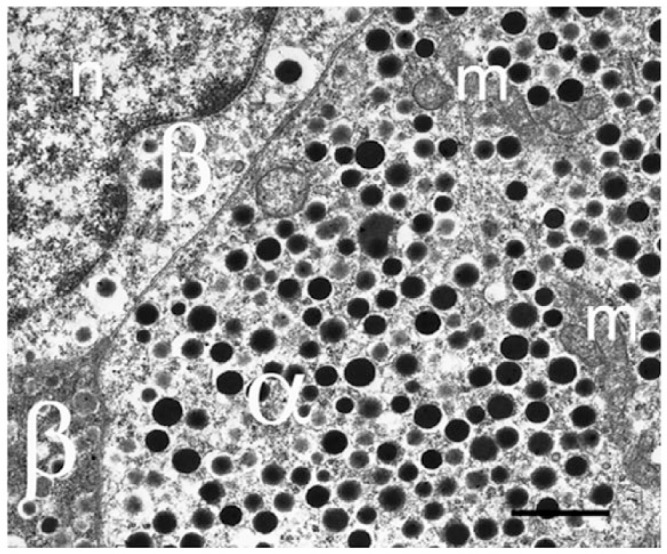
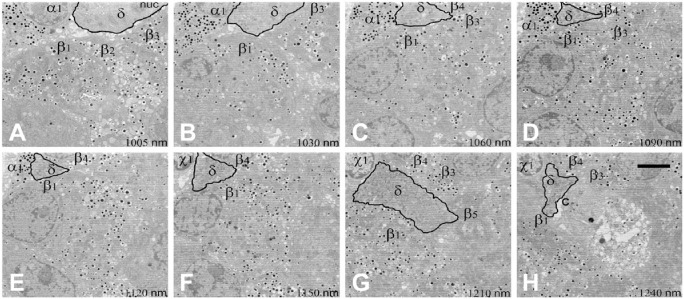
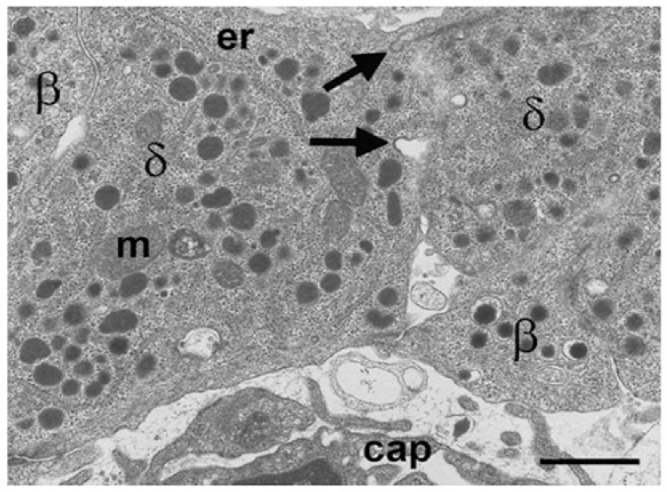
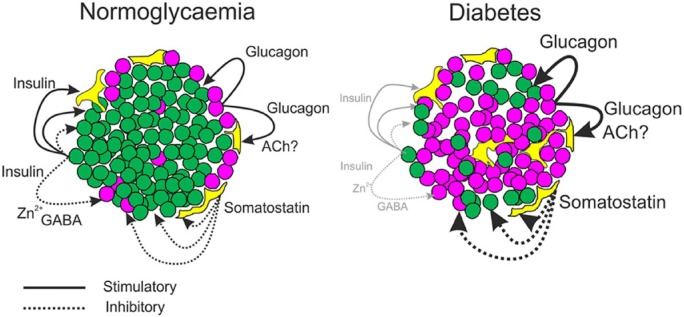
Islet non-β-cells, the α- δ- and pancreatic polypeptide cells (PP-cells), are important components of islet architecture and intercellular communication. In α-cells, glucagon is found in electron-dense granules; granule exocytosis is calcium-dependent via P/Q-type Ca(2+)-channels, which may be clustered at designated cell membrane sites. Somatostatin-containing δ-cells are neuron-like, creating a network for intra-islet communication. Somatostatin 1-28 and 1-14 have a short bioactive half-life, suggesting inhibitory action via paracrine signaling. PP-cells are the most infrequent islet cell type. The embryologically separate ventral pancreas anlage contains PP-rich islets that are morphologically diffuse and α-cell deficient. Tissue samples taken from the head region are unlikely to be representative of the whole pancreas. PP has anorexic effects on gastro-intestinal function and alters insulin and glucagon secretion. Islet architecture is disrupted in rodent diabetic models, diabetic primates and human Type 1 and Type 2 diabetes, with an increased α-cell population and relocation of non-β-cells to central areas of the islet. In diabetes, the transdifferentiation of non-β-cells, with changes in hormone content, suggests plasticity of islet cells but cellular function may be compromised. Understanding how diabetes-related disordered islet structure influences intra-islet cellular communication could clarify how non-β-cells contribute to the control of islet function.
Keywords: PP; communication; exocytosis; glucagon; granule; insulin; intra-islet signaling; non-β-cell; paracrine; somatostatin.
© The Author(s) 2015.
Conflict of interest statement
Figures
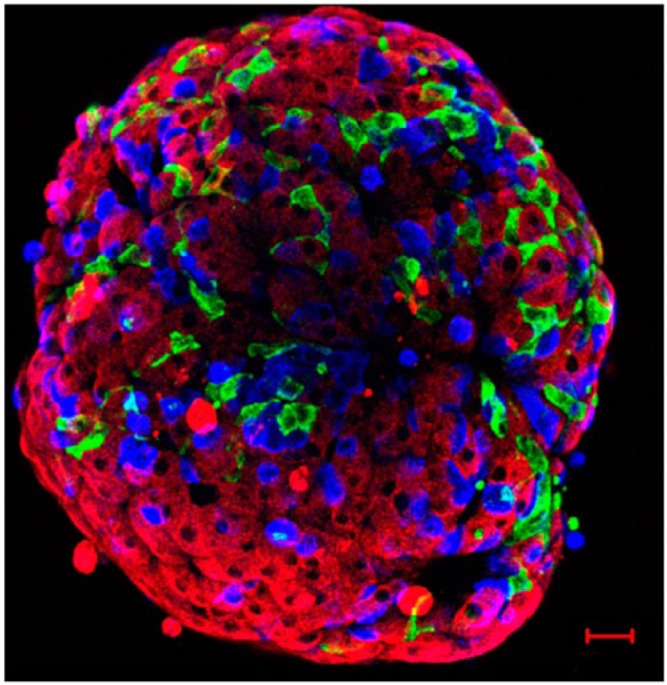
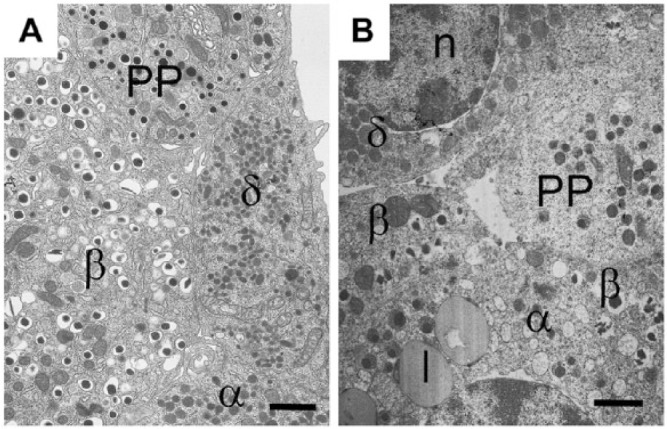
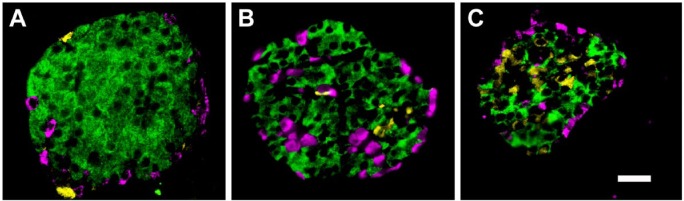
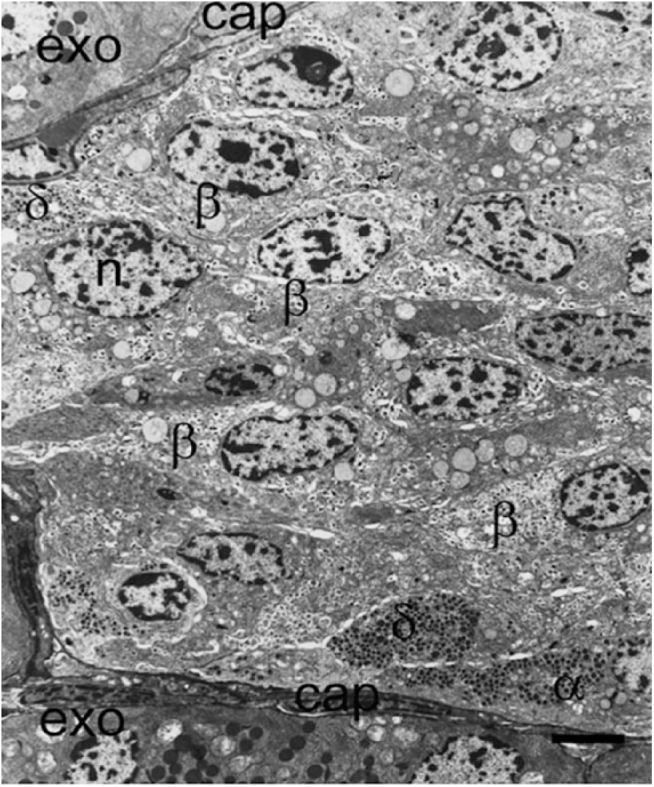
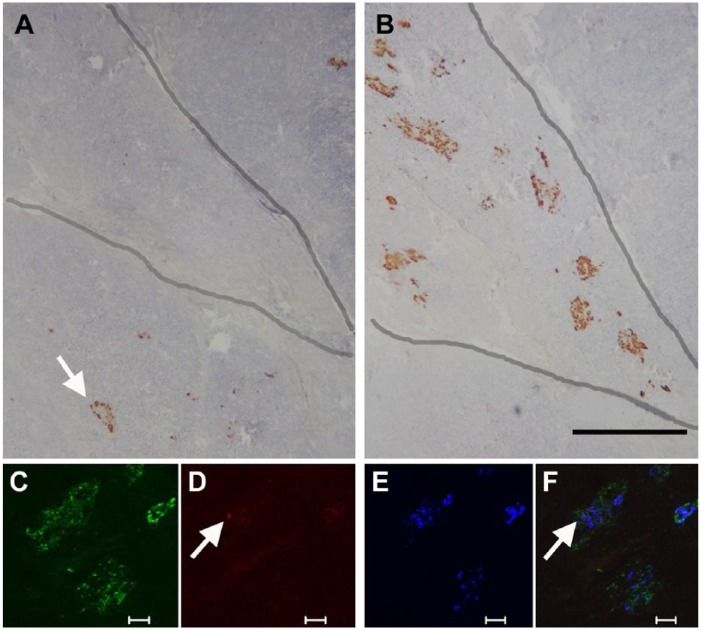
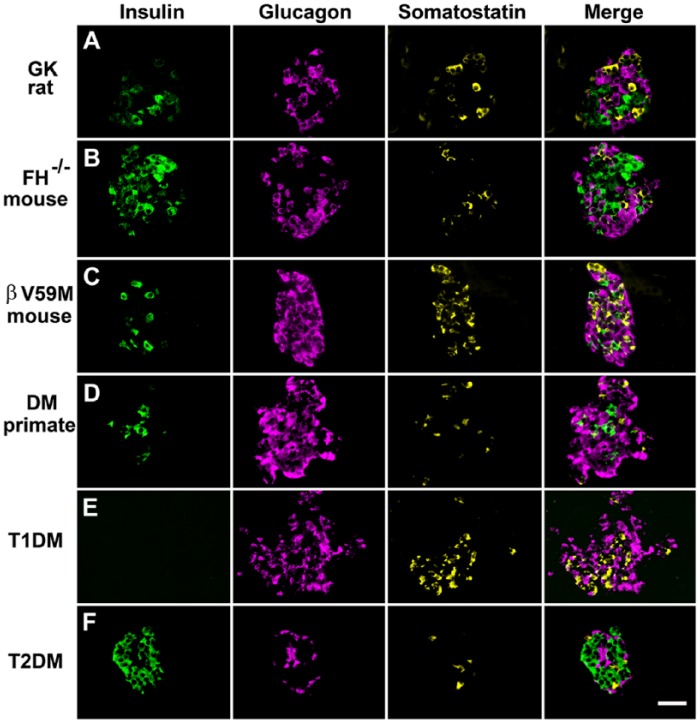
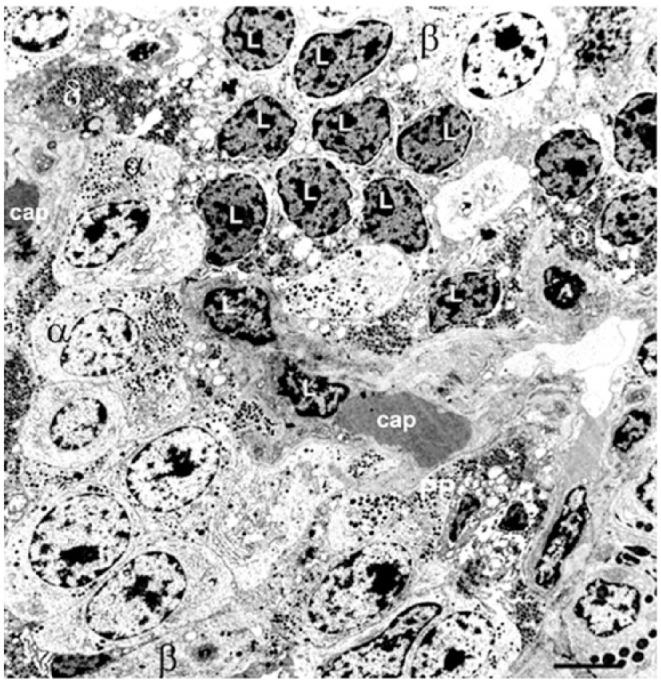
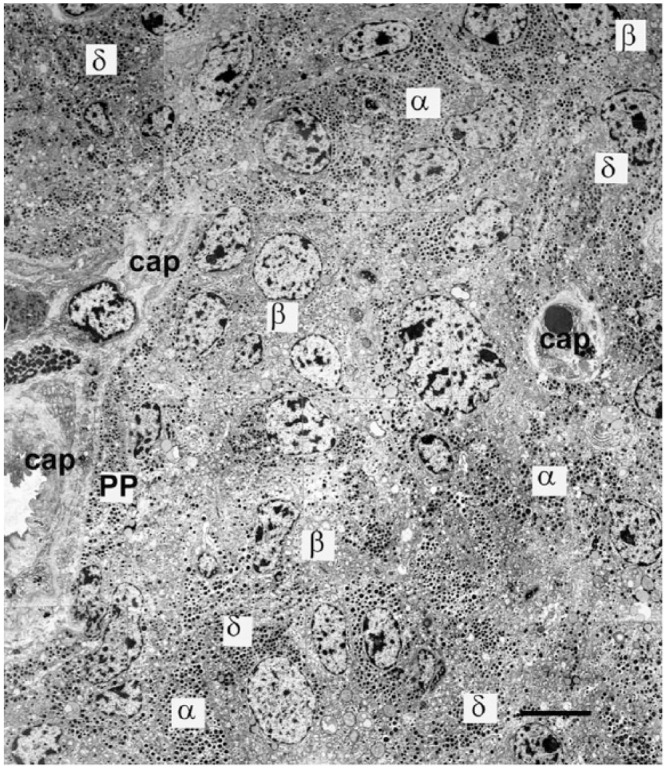
References
-
- Aragon F, Karaca M, Novials A, Maldonado R, Maechler P, Rubi B. (2015). Pancreatic polypeptide regulates glucagon release through PPYR1 receptors expressed in mouse and human alpha-cells. Biochim Biophys Acta 1850:343-351. - PubMed
-
- Arimura A, Sato H, Dupont A, Nishi N, Schally AV. (1975). Somatostatin: abundance of immunoreactive hormone in rat stomach and pancreas. Science 189:1007-1009. - PubMed
-
- Barg S, Galvanovskis J, Gopel SO, Rorsman P, Eliasson L. (2000). Tight coupling between electrical activity and exocytosis in mouse glucagon-secreting alpha-cells. Diabetes 49:1500-1510. - PubMed
-
- Baron AD, Schaeffer L, Shragg P, Kolterman OG. (1987). Role of hyperglucagonemia in maintenance of increased rates of hepatic glucose output in type II diabetics. Diabetes 36:274-283. - PubMed
-
- Baskin DG, Gorray KC, Fujimoto WY. (1984). Immunocytochemical identification of cells containing insulin, glucagon, somatostatin, and pancreatic polypeptide in the islets of Langerhans of the guinea pig pancreas with light and electron microscopy. Anat Rec 208:567-578. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
